|
Desert Varnish And Lichen Crust
Desert Varnish & Lichen Crust
Microscopic Organisms Color Rocks & Boulders
|
Major Topics To Be Discussed:
|
|
Images Of Desert Varnish And Rock Lichens:
|
Introduction
|
Rugged mountain peaks and sun-baked
boulders throughout the arid southwest are often colored in
beautiful shades of orange, green, yellow and gray. At first
glance the colorful coatings resemble a layer of paint, but close
examination reveals that this unusual phenomenon is caused by a
thin layer of microscopic organisms. The organisms include colonies of bacteria called "desert varnish" (Kingdom Monera), and colonies of symbiotic algae and fungi called lichens (Kingdoms Protista and Fungi). These miniature rock
dwellers have survived for countless centuries in some of the
most seemingly inhospitable environments on earth, and may
represent some of the oldest living colonial life forms.
Symbiogenesis: New Species
Resulting From Genomic Mergers
Long-term stable symbiotic relationships between two or more organisms may lead to evolutionary change. This phenomenon, known as symbiogenesis, is described in the book entitled Acquiring Genomes: A Theory of the Origins of Species by L. Margulis and D. Sagan (2002). According to the authors, genomic mergers are a major source of genetic variability leading to the evolution of species. Instead of relying on the hit-or-miss method of random mutations, symbiogenesis provides advantageous genetic combinations through the fusion of entire genomes from two or more organisms. This phenomenon may have been a major factor in the evolution of land plants from lichen-like ancestors.
Endosymbiont Hypothesis: The Evolution Of Chloroplasts
The arise of photosynthetic organelles called chloroplasts represented a major step in the evolution and diversification of plant life on earth. This event had a significant effect on the evolution of animal life that depended on the plants for food, either directly in the case of herbivores, or indirectly in the case of carnivores. Chloroplasts exhibit remarkable similarities with cells of cyanobacteria, and may have shared a common prokaryotic ancestry. Indeed, the outer membrane structure and circular DNA molecules of chloroplasts and mitochondria are very similar to individual prokaroytic cells. According to the endosymbiont hypothesis (or endosymbiont theory for those who are less skeptical), ancient photosynthetic prokaryotic cells became incorporated within the cells of algae or ancestral plants, forming stable mutualistic symbionts known as chloroplasts. Mitochondria may have had a similar origin. Without chloroplasts and oxygen-producing photosynthesis, the amazing diversity of today's plants and animals would not have evolved.
According to Margulis and Sagan (2002), there is even a green photosynthetic animal named Elysia viridis, a minute slug (saccoglossan opisthobranch) that never feeds throughout its adult life. Instead, it obtains carbohydrate-rich molecules by bathing in the sunlight. This slug evolved from algae-eating ancestors, only the algal cells entered the slug's tissue and remained their as photobionts (photosynthetic symbionts). According to some invertebrate zoology textbooks, the chloroplasts from algae cells are sucked into the slug's gut and incorporated within digestive gland cells where photosynthesis occurs.
There are numerous examples of plants and animals that contain microbial symbionts within their tissues, including bacteria, cyanobacteria, protozoans and algal cells. Cycads, water ferns (Azolla), legumes and the tropical flowering plant Gunnera contain nitrogen-fixing bacteria in their tissues; sea anemones and corals contain photosynthertic unicellular algae (zooanthellae and zoochlorellae); the rumen of cattle contain cellulose-digesting bacteria; termite guts contain flagellated protists which contain wood-digesting bacteria; and human intestines contain bacteria that produce essential B vitamins.
Lichens are one of the best examples of symbiogenesis involving the fusion of algal and fungal genomes (kingdoms Protista and Fungi). Some lichens include the genome of a third kingdom Monera because they contain prokaryotic cells of cyanobacteria. In the case of lichens, this genomic merger has enabled these organisms to survive in some of the most inhospitable environments on earth, where neither symbiont could survive on its own. In fact, lichens are an excellent example of synergism because the whole is truly greater than the sum of its parts. The algal and fungal components develop into a unique body form with morphological features quite different from either symbiont.

|
British soldiers (Cladonia cristatella), a soil lichen with upright podetia bearing bright red apothecia at the tips. At the bottom of the centrifuge tube (left), the fungal component of this lichen (also named C. cristatella) has grown into a white, amorphous blob without its algal symbiont. In the right test tube, the algal symbiont (named Trebouxia erici) has grown into a mass of bright green cells. Only when these two symbionts form the "marriage" known as lichen is the unique structure of "British soldiers" formed. In true synergistic fashion, the lichen is truly more than the sum of its parts. For example, the podetium is a unique lichen structure that is not found in the algae or fungi.
|

|
Gel electrophoresis comparing DNA banding from a lichen Cladonia cristatella (lane L) with its symbiotic components, the photobiont alga Trebouxia erici (lane A) and the mycobiont fungus (lane F) which is also named Cladonia cristatella. Molecular weights of the different DNA fragments are shown in the far left column (S). The autotrophic alga is photosynthetic and provides carbohydrate nutrition for the heterotrophic fungus. The lichen DNA (lane L) shows some banding that is different from the patterns of its two symbionts. In true synergistic fashion, the lichen is truly more than the sum of its parts. [Gels courtesy of J.L. Platt, CSUSM, San Marcos, California.]
|
Lichens & The Evolution of Land Plants
Lichenization, the process by which fungal hyphae and algal cells literally grow together to form a mutualistic association, may help to explain the remarkable evolution of vascular plants on earth. Most textbooks of general botany suggest that land plants evolved from ancestral green algae. However, some authorities believe that vascular plants are far more than simple extensions of green algae. They are comparatively too complex, diversified too quickly, and contain numerous fungus-like cells. In his fascinating article "Are Vascular Plants Inside-Out Lichens?" (Ecology 69 (1): 17-23, 1988), Peter Atsatt of the University of California, Irvine discusses some of the evidence supporting a lichenized ancestor to vascular plants. According to Dr. Atsatt, the ancestral lichenization resulted in a "reverse-phase lichen" with a dominant algal component containing an endophytic, mineral scavenging fungus similar to extant mycorrhizal associations. Nuclear fusions between the fungal and algal cells resulted in hybrid nuclei containing the traits of both parents. In true synergistic fashion, this dual genome gave rise to a plant body composed of a mosaic of alga-like photosynthetic cells interspersed with specialized fungus-like cells.
Probably the most difficult concept for skeptical botanists to accept is the fungal ancestry in today's vascular plants. Dr. Atsatt discusses several types of cells and tissues in vascular plants which resemble fungal hyphae, including pollen tubes, vascular (xylem) tissue, laticifers, and haustoria. Pollen tubes not only resemble the growth of fungal hyphae, but in Pinus, cycads, and Ginkgo they are branched and actually absorb nutrients from the "host's" megasporangium. The latex-producing laticifers found in many members of the Euphorbiaceae, Asclepiadaceae and other dicotyledonous families are very similar to fungal hyphae. Nonarticulated laticifers are elongate, multinucleate cellular tubes that grow throughout the plant body of these families. Some endophytic parasitic flowering plants, such as certain dwarf mistletoes and the remarkable Pilostyles thurberi of the Colorado Desert, live completely within the host tissues and only emerge from their host to produce flowers. The vascular tissue of these endoparasites literally permeate the host tissues and truly resemble fungal hyphae. The absorptive haustorial organs of many parasitic flowering plants which penetrate the host tissue are also very reminiscent of fungal hyphae.
Additional evidence of fungal ancestry in land plants are minute bodies called "mycosomes." Dr. Atsatt has discovered these minute fungal propagules inside the plastids of algae, primitive and higher plants, including mosses (Polytrichum), club-mosses (Lycopodium), cycads (Cycas revoluta), Ginkgo biloba, conifers (Sequoia sempervirens) and mormon tea (Ephedra), parasitic flowering plants (Phoradendron,Cuscuta, Cassytha and Pilostyles), marine flowering plants (Zostera) and Indian paintbrush (Castilleja stenantha). Mycosomes are apparently widespread in plant and algal cells, and are produced by both ascomycete and basidiomycete fungi. [See Atsatt, P.R. 2003. "Fungus Propagules in Plastids: The Mycosome Hypthesis." International Microbiology 6: 17-27.] Dr. Atsatt defines mycosomes as "wall-deficient reproductive propagules (diameter ca 0.1-1.0 μm) that develop into larger forms: wall-less cells (protoplasts), yeast cells, or membrane-bounded thylakoid-containing spherules in which mycosomes reproduce."
Although there is ample fossil evidence suggesting that algae and fungi lived over 500 million years ago, there is little fossil evidence of true ancestral lichens from that era. However, a startling new hypothesis from Gregory Retallack of the University of Oregon may shed some light on the existence of Precambrian lichens. Since their discovery in southern Australia in the late 1950s, the fossil remains of the Ediacaran biota have puzzled paleobiologists. These strange, flattened creatures lived about 600 million years ago presumedly at the bottom of shallow coastal seas. They have been classified in several primitive animal phyla, from jellyfish, echinoderms and worm-like animals to large alga-like protists, and may have been ancestral to other animal phyla. A number of paleontologists refer to these organisms as "Vendobionta," and regard them as an extinct early experiment in the evolution of life. By about 530 million years ago they were all replaced by shelled Cambrian animals. But according to Dr. Retallack, these bizarre creatures may have been ancient lichens. In his fascinating article "Were the Ediacaran Fossils Lichens?" (Paleobiology 20 (4): 523-544, 1994), Dr. Retallack eloquently discusses the evidence supporting his lichen hypothesis. Comparing their thickness to that of much younger tree-trunk fossils, he concludes that the fossils resisted compaction after burial almost as well as sturdy logs. Their sturdiness, large size (up to one meter across), lack of any mouth, digestive cavity or musculature, and evidence from their microscopic examination all suggest to Retallack that Ediacaran fossils were lichens. The presumed marine habitats of Ediacaran fossils is not crucial to their interpretation as lichens, because rock lichens live in the sea and on land. If one can hypothesize that at least some of the Ediacarans may be ancestral to certain animal groups, then perhaps a lichen-like Ediacaran ancestor gave rise to vascular plants!
According to Blair Hedges of Pennylvania State University (Science, August 2001), aquatic fungi evolved into a terrestrial form about 1.3 billion years ago. These early fungal forms were actually lichens because they formed a symbiotic relationship with primitive aquatic green algae. The early land surface of the Earth at this time contained numerous colorful rock lichens. The bright pigments served to reduce the harmful effects of ultraviolet radiation in a primitive atmosphere. Evidence from mutation rates in 119 genes common to living fungi and plants, indicates that ancient moss-like land plants appeared about 700 million years ago.
Lichens are far more than mere biological curiosities. They are very successful and unique life forms that may hold the secrets to complex evolutionary processes, cell differentiation and gene expression in vascular plants.
|
Wayne's Word Hyperlinks About
Symbiosis
|
Desert Varnish On Rocks And Boulders
One of the most remarkable
biogeochemical phenomena in arid desert regions of the world is
desert varnish. Although it may be only a hundredth of a
millimeter in thickness, desert varnish often colors entire
desert mountain ranges black or reddish brown. Desert varnish is
a thin coating (patina) of manganese, iron and clays on the
surface of sun-baked boulders. According to Ronald I. Dorn and
Theodore M. Oberlander (Science Volume 213, 1981), desert
varnish is formed by colonies of microscopic bacteria living on
the rock surface for thousands of years. The bacteria absorb
trace amounts of manganese and iron from the atmosphere and
precipitate it as a black layer of manganese oxide or reddish
iron oxide on the rock surfaces. This thin layer also includes
cemented clay particles which help to shield the bacteria against
desiccation, extreme heat and intense solar radiation.
|
In the Anza-Borrego Desert of southern California, the sun-baked boulders are devoid of lichens. Instead, the rocks are coated with "desert varnish," a reddish layer of clay and iron oxide precipitated by remarkable bacteria. This large boulder has split apart revealing the lighter granodiorite beneath.
|
Several genera of bacteria are
known to produce desert varnish, including Metallogenium
and Pedomicrobium. They consist of minute spherical, rod-
shaped or pear-shaped cells only 0.4 to 2 micrometers long, with
peculiar cellular extensions. In fact, the individual cells are
smaller than human red blood cells which are about 7.5
micrometers in diameter. Because of the radiating filaments from
individual cells and colonies, they are called appendaged
bacteria. All living systems require the vital energy molecule
ATP (adenosine triphosphate) in order to function. In our cells
ATP is constantly produced within minute bodies called
mitochondria. As electrons flow along the membranes of our
mitochondria, molecules of ATP are generated. The electrons come
from the breakdown (oxidation) of glucose from our diet.
Although varnish bacteria do not have mitochondria, they do have
a similar inner membrane structure through which electrons flow
to generate ATP. However, in varnish bacteria the electrons come
from the oxidation of manganese and iron rather than glucose.
Herein lies the marvelous adaptive advantage for producing a
layer of black and red varnish on desert boulders.

Manganese varnish formation. Modified from Dorn & Oberlander,
1982.
Reddish iron varnish develops in a similar oxidation reaction.
|
Varnish bacteria thrive on smooth
rock surfaces in arid climates. According to Ronald Dorn,
perhaps 10,000 years are required for a complete varnish coating
to form in the deserts of the southwestern United States. In
fact, dating of varnished surfaces is of enormous importance to
the study of desert landforms and to the study of early humans in
America, since many artifacts lying on the ground become coated
with desert varnish. Boulders of the Anza-Borrego Desert region
are covered with a reddish-brown iron oxide, while boulders in
parts of Owens Valley are blackened by a manganese oxide varnish.

|
The thin layer of reddish iron oxide varnish on this rock surface has been etched to reveal the lighter granodiorite beneath. Several Indian tribes utilized desert varnish to create their marvelous petroglyphs.
|
In the Alabama Hills near Lone Pine, the rocks are so black that they resemble basalt; however, if you scratch through the varnish
layer, the light-colored granite is exposed. For thousands of
years native Indians have used desert varnish for their rock
carvings (called petroglyphs). Throughout northern Owens Valley,
there are acres of elaborate petroglyphs carved into desert
varnish and Bishop tuff, including spirals, circles, wavy lines,
footprints, men, deer and desert bighorn sheep. It is
fascinating to speculate on the origin and meaning of all these
carvings.

|
This varnish-coated rock (with black layer of manganese oxide) in the Alabama Hills near Lone Pine, California has broken away revealing the lighter granitic core.
|
Lichen Crust On Rocks And Boulders
Without a doubt, the most colorful
coatings on rocks are produced by lichens, a remarkable symbiotic
relationship between microscopic algal cells and fungal
filaments. Although lichens can also withstand extreme
environmental conditions, they generally cannot survive as well
on the dry, sun-baked boulders where desert varnish microbes flourish.
Rock lichens come in a variety of bright colors, from red, orange
and yellow to brilliant shades of green. There are hundreds of
species in the southwestern United States, including leafy forms
and low-growing crustose species resembling a thick layer of
paint.
|
This basalt outcrop on the Santa Rosa Plateau of southern California is covered by a dense crust of four colorful lichen species, including orange Caloplaca, yellow Candelaria & Candelariella and gray Xanthoparmelia.
|
The lichen body is composed of algal cells living inside a compact mass of fungal tissue. The algae are photosynthetic
and provide the fungus with carbohydrate nutrients. Three genera of unicellular green algae (Trebouxia, Pseudotrebouxia and Trentepohlia) are involved in the majority of lichens. Two genera of filamentous cyanobacteria (Nostoc and Scytonema) are also found in many species of lichens. In fact, some remarkable lichens contain two symbionts, an alga and a cyanobacterium.
|
Placopsis gelida: A striking North American crustose lichen containing two autotrophic symbionts (algae and cyanobacteria). The pinkish bodies in the center (called cephalodia) contain nitrogen-fixing cyanobacteria.
|
The delicate
algal and bacterial cells also gain mechanical protection from hostile climatic
conditions by being tightly enveloped in a dense meshwork of
fungal filaments. This is especially true on arid boulders where
neither partner could survive on its on. Indeed the relationship
is a kind of marriage where each member depends on the other for
its survival. It wasn't until 1867 that the dual symbiotic
nature of lichens was described by the Swiss botanist Simon
Schwendener. One of the staunch proponents of the alga/fungus
marriage hypothesis was the excellent naturalist and scientific
illustrator of this time by the name of Beatrix Potter--who
latter became a famous author of children's stories.

|
A granite boulder in the Sierra Nevada of California covered with colonies of map lichen (Rhizocarpon geographicum). This species of crustose lichen is appropriately named because the colonies resemble continents on the earth.
|
|
|
Left: Several species of crustose lichens on the surface of Santiago Peak Volcanic rock in the Coast Ranges of southern California. The lichens include lemon-yellow Acarospora socialis, brown A. bullata, and gray Dimelaena radiata. The Acarospora doesn't have the marginally lobate thallus of Pleopsidium flavum and P. chlorophanum. The brown crostose lichen A. bullata is similar to the common species A. fuscata. Note the marginally lobate thallus of D. radiata. Right: Close-up view of the areolate thallus of Acarospora reveals a minute tardigrade nestled in a crack in the thallus.
|
|
Lichens also have diminutive animals living on their surface. The most remarkable are tardigrades, minute creatures that belong to their own Phylum Tardigrada. Tardigrades are only about .25 mm long (1/100 of an inch). Although they are fairly common on mosses and certain lichens, they are rarely seen. They are called "water bears" because of their peculiar fat body with stubby legs bearing claws at the tips. When lichens become desiccated and dormant during drought periods, the tardigrades also go into a stage of suspended animation known as anabiosis. They pull in their legs, lose water, and roll up into a ball. When moisture is available again, the tardigrades swell and become active within a few hours. There are records of tardigrades emerging from a state of anabiosis lasting seven years.

|
Minute tardigrades (Phylum Tardigrada) are only about 0.25 mm long (1/100 of an inch). Although they are fairly common on mosses and lichens, they are rarely seen. They are called "water bears" because of their fat body with stubby legs bearing claws at the tips. The following illustration was enhanced from this photograph to show the legs and claws.
|

|
An illustration of a tardigrade that was enhanced from the above photograph to show the legs and claws.
|
Microscopic examination reveals that lichens are really fungi containing photosynthetic algal cells. In fact, they are called "lichenized fungi" by mycologists (people who study fungi). The lichen body is called a thallus, and is composed of intertwined fungal filaments. Lichens are conveniently grouped into 3 characteristic growth forms. Foliose lichens have a leaf-like or lobed thallus, loosely attached to the substrate by root-like rhizines or by a central stalk-like umbilicus. Fruticose lichens have an intricately branched upright or pendulous thallus. Crustose lichens are low-growing, with the entire thallus firmly attached to the rock or other substrate. The thallus of crustose lichens may be squamulose (composed of crowded, overlapping scales) or areolate (cracked into numerous angular sections like a dried lake bed).
A remarkable biochemical phenomenon resulting from the alga-fungus union in lichens is the production of lichen acids, unique phenolic compounds produced by no other organisms. In fact, they are not even produced by the separate fungal or algal symbionts, only by the combined lichen. Lichen acids have the unusual property of changing colors under different pH conditions. In fact, the litmus dye that turns red in acid solutions and blue in alkaline solutions was originally derived from the fruticose lichen Roccella. The color change under different pH conditions is caused by changes in the electron configuration (resonance) of the molecule. This affects absorption and reflection of light; hence, the vivid color changes.
T he presence of certain lichen acids is used to identify some species. A drop of sodium hydroxide or lye (NaOH) and a drop of sodium hypochlorite or household bleach (NaClO) is added to the exposed lichen tissue (cortex and medulla) to see if it turns yellow, purple or red. [The cortex is the outermost layer of the thallus composed of compressed hyphal cells. The medulla is the innermost layer of the thallus composed of loosely interwoven hyphae and algal cells.] Since potassium hydroxide (KOH) rather than NaOH has been commonly used on lichens, the hydroxide test is abbreviated as K. The hypochlorite test is abbreviated as C. K+ yellow indicates a positive reaction for atranorin (cortex), alectorialic acid (medulla) and stictic acid (medulla). K+ yellow turning red indicates a positive reaction for norstictic acid and salazinic acid (medulla). C+ red indicates a positive reaction for gyrophoric acid, lecanoric acid and olivetoric acid (medulla). The orange cortical pigment parietin in Caloplaca, Teloschistes and Xanthoria has a K+ reaction of deep purple. Of course, K- and C- indicates a negative reaction with no color change.
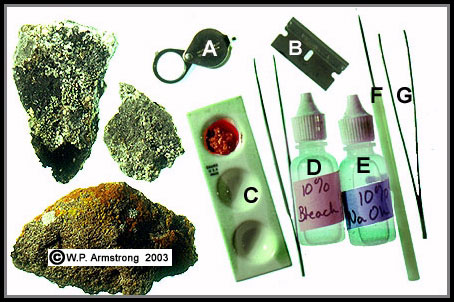
|
Tools for the field identification of lichens: A. 20x hand lens, B. single-edge razor blade, C. depression slide for testing lichen sample with NaClO and NaOH, D. 10% sodium hypochlorite or bleach (NaClO), E. 10% sodium hydroxide or lye (NaOH), F. needle probe and F. sharp-pointed forceps.
|
Lichen acids were the source of important dyes for cotton and wool in medieval Europe. Two purple and red dyes, orchil (sometimes spelled archil) and cudbear, were obtained from the lichens Roccella and Ochrolechia. Lichen dyes were dissolved in human urine, and the yarns were immersed in this mixture. Ammonia salts in the urine functioned as mordants to make the dyes permanent. Although most of the lichen dye industry was replaced by cheaper aniline dyes from coal tar, some lichen dyes are still used today. A brownish dye from the foliose lichen Parmelia omphalodes is used on hand-woven Harris tweeds from the Outer Hebrides. Orcein, a purple-red chromosomal stain found in every microbiology laboratory is derived from Roccella tinctoria. Pine lichen (Letharia vulpina), a beautiful chartreuse fruticose lichen that grows on the bark of pines and fir throughout the mountains of the Pacific United States, contains a mildly toxic yellow dye called vulpinic acid. The striking canary-yellow porcupine quills woven into the baskets of Klamoth and Yurok Indians were dyed with this lichen.

|
The purple-red stain orcein is derived from the lichen Roccella tinctoria. In fact, the washcloth in photo was dyed with orcein. Without a mordant, such as urea, the colors will readily wash out of the cloth.
|
Some lichens contain various phenolic acids and essential oils that produce fragrant odors in scented soaps and help fix the aroma of fine perfumes. For centuries a lovely fruticose lichen called oak moss (Evernia prunastri) has been collected in Europe for making perfume.Through a complex process of solvent extraction and distillation, oak moss has become an important ingredient in the manufacture of perfumes and high-quality cosmetics. This remarkable lichen occurs in California, but air pollution has eliminated it throughout most of its former range in southern California. Oak moss still clings to the branches of ponderosa pines on Palomar Mountain in San Diego County.

|
An assortment of fruticose and foliose bark lichens from white fir, ponderosa pine (Pinus ponderosa) and incense cedar (Calocedrus decurrens) on Palomar Mountain in San Diego County: A. Oak moss (Evernia prunastri), the same species collected in Europe for a delicate perfume; B. pine lichen (Letharia vulpina); C. Old man's beard (Usnea); [Thallus branches of the latter species have a central cord, thus ruling out the similar lichen Alectoria.] D. Platismatia glauca (technically a foliose lichen); E. Hypogymnia imshaugii (technically a foliose lichen); F. Old man's beard (Bryoria fremontii).
|
Even without magnification, one of the most visible fungal characteristics of a lichen is the cup-shaped, spore-bearing structure called an apothecium. This structure is found in many related fungi belonging to the Class Ascomycetes, and is produced through a complex sexual cycle. The cup-shaped apothecia of some lichens resemble those of non-lichenized fungi, with a rim or margin of fungal hyphae (called a proper exciple) that is usually the same color as the cup. Many lichens develop a second (outer) rim the same color as the thallus. This uniquely lichen structure (called a thalline exciple) contains a layer of algal cells. Usually only one type of rim is visible under a hand lens, but some lichens have both types. These terms are important when keying out certain lichens, especially crustose species.

|
Cup-shaped apothecia of Xanthoparmelia cumberlandia, a common foliose rock lichen throughout San Diego County. The apothecium is lined with microscopic saclike structures called asci, each bearing eight ascospores. This reproductive structure is produced by the fungal component (symbiont) of the lichen. Nonlichenized fungi (without the algal symbiont) in the class Ascomycetes also produce cup-shaped apothecia. Because of their distinct apothecia, members of the class Ascomycetes are often referred to as cup fungi.
|
The thallus of some lichens is dotted with black pits called pycnidia, which are lined with minute, asexual conidia. These bacilliform spores (also called pycnidiospores) are strikingly similar in appearance to the male spermatia produced by certain non-lichenized fungi such as wheat rust. The following photo shows the common foliose bark lichen Hypogymnia imshaugii that grows on conifer branches in the mountains of San Diego County.

|
The foliose bark lichen Hypogymnia imshaugii, showing numerous, minute black pits called pycnidia on the surface of the grayish-green thallus.
|
Presumably the fungal spores produced in apothecia may encounter algal cells on a rock or branch and begin a new career as conjugal partners, in a union observed in the laboratory but rarely seen in nature. However, lichens have evolved a much more effective method of asexual reproduction and dispersal through the production of soredia and isidia. Soredia are microscopic particles containing several algal cells enveloped by fungal hyphae. They are produced in pustules on the surface of the lichen called soralia. Like dust particles, the soredia become airborne and are probably responsible for the worldwide distribution of some lichens. Isidia are pimplelike protuberances containing both algae and fungi that readily disperse to new locations.
The following illustration shows a typical foliose lichen in cross section. The lower surface has root-like extensions called rhizines. The upper side has a cup-shaped apothecium lined on the inside with numerous sac-like structures called asci. Each ascus produces 8 ascopores which escape from the cup by wind and rain. The upper side also has a blister-like pustule called a soralium bearing numerous particles called soredia. Each soredium is composed of several green algal cells enveloped by fungal filaments (hyphae). It is interesting to note that some lichen characteristics, such as soredia, are not produced by typical fungi. They are only produced by the algal-fungus association through some unknown genetic mechanism.
|

|
Cross section through the thallus of a typical foliose lichen.
|

|
Left: Flavoparmelia caperata, a foliose bark lichen showing several soralia erupting from the upper surface of the thallus. Center: Microscopic view of soredia from the common chaparral lichen F. caperata. Each spherical soredium contains several green algal cells enveloped by filamentous fungal hyphae. Like dust particles, the soredia are carried by the wind to different locations where they develop into new lichens. Right: Closeup view of the surface of the foliose rock lichen Xanthoparmelia mexicana showing pimplelike projections called isidia. The isidia contain algae and fungi and readily become detached and dispersed to new locations.
|
|
Some crustose lichens produce a flask-shaped perithecium rather than a cup-shaped apothecium. Without magnification the perithecia appear like tiny pimples on the lichen surface. Under high magnification the perithecium appears like a miniature flask or volcano, open at one end and lined on the inside with numerous asci, each bearing 8 ascospores. The ascospores escape through the pore at the top of the perithecium. The following illustration shows a cross section of a perithecium lined on the inside with numerous spore-bearing asci.

|
Cross section through the perithecium of a crustose lichen. For close-up photos of perithecia, see the following freshwater and marine lichens.
|

|
Microscopic view inside the perithecium of Verrucaria viridula showing several saclike asci and a cluster of eight ascospores. The colorless spores are nonseptate, and not divided into sections as in the following crustose lichen Staurothele.
|

|
Species of Verrucaria and Staurothele grow on moist rocks and boulders and superficially resemble each other. They both have an areolate thallus that resembles a thin layer of dried, cracked mud, and they both have pimplelike perithecia; however, their spores are very different. Verrucaria has eight colorless, nonseptate spores per ascus. Staurothele has two spores per ascus. The two large spores of Staurothele are reticulately sculptured (muriform). Each spore is partitioned into sections like the squares of a Hershey's® chocolate bar.
|
|

|
Left: The crustose lichen Staurothele fissa covers the boulders along Fern Creek on the eastern side of the Sierra Nevada. Center: The cracked, areolate thallus of Staurothele fissa superficially resembles a layer of dried mud, but close examination reveals minute, pimplelike fungal fruiting bodies (perithecia), each with a terminal pore. Right: Microscopic view (400x) of the ascospores of Staurothele fissa. The spores are reticulately sculptured (muriform) like the squares of a chocolate bar. Staurothele has only two spores per ascus.
|
Crustose Marine Lichens Of The Pacific Coast

|
The Verrucaria maura zone (v.m.) on the coast of central Oregon at low tide. This crustose marine lichen forms a distinct dark band above the barnacle zone along the rocky shores of Oregon and Washington. It is sometimes mistaken for hardened oil that has washed ashore.
|
|
Close-up view of the crustose, areolate lichen Verrucaria maura. The pimplelike structures are perithecia, each with a minute apical pore. This marine lichen grows on intertidal rocks along the wave-battered, rocky shores of the Pacific United States and Canada. Related species cover boulders and rocky outcrops throughout North America and other continents of the world.
|

|
Caloplaca coralloides, a colorful marine lichen that grows in the intertidal spray zone along the rocky coast of Oregon. Unlike the majority of crustose species of Caloplaca, this interesting lichen is fruticose.
|
|
According to Lichen Flora of the Greater Sonoran Desert Region (Vol 1) by T. H. Nash et al. (2002), the following species Pyrenocollema halodytes is now listed as Collemopsidium sublitoralis. It has been verified from Cabrillo National Monument by K. Knudsen (personal communication, 2008).
|

|
Left: The rocky intertidal zone of Point Loma (San Diego County): Home of the interesting marine lichen Pyrenocollema halodytes. Right: The microscopic, conical, fungal fruiting bodies (perithecia) are scattered over the surface of boulders. Each perithecium has a terminal opening or pore and resembles a microscopic volcano.
|
|
Left: Close-up view of an intertidal rock in southern California showing black patches of the marine crustose lichen Pyrenocollema halodytes (red arrow). The head of a sewing needle is shown as a size relationship. Right: Microscopic view (100x) of the perithecium of the marine lichen Pyrenocollema halodytes (red arrow). The perithecium is minute, only 0.25 mm in diameter. The thallus of this species is often endolithic, growing below the surface of rocks, sandstone and the calcareous shells of gastropods and acorn barnacles. The colorless spores are 1-septate (2-celled), unlike the simple, non-septate spores of Verrucaria.
|

|
Left: Microscopic view of the hymenial layer from a perithecium of Pyrenocollema halodytes. The ascospores are 1-septate (2-celled), with one cell slightly larger. Several saclike asci are shown in the photo, one of which has split open releasing ascospores. Right: Microscopic view of the photobiont of Pyrenocollema halodytes showing several filaments of cyanobacteria. The larger, oval cell is a heterocyst, the site of nitrogen fixation. In addition to producing carbohydrate sugars for its fungal partner, this blue-green photobiont also converts inert atmospheric nitrogen into a usable form called ammonia.
|

|
Left: The tiny pits (red circles) on these acorn barnacles contain the fungal fruiting bodies (perithecia) of the marine lichen Pyrenocollema halodytes. Since barnacles commonly grow on the backs of gray whales, it is quite possible that this interesting endolithic lichen may also live in barnacles that, in turn, live on whales. Right: Another marine lichen with minute, pimplelike perithecia (red circles) grows on the wave-battered sandstone shoreline of southern California with limpets, chitons and barnacles. The following microscopic image clearly shows that it is not Pyrenocollema halodytes.
|

|
Microscopic view of several spores and asci from a pyrenocarpus intertidal marine lichen. Each ascus contains eight colorless, nonseptate ascospres. This is clearly not Pyrenocollema halodytes because Pyrenocollema has 1-septate spores and a cyanobacterial photobiont. The thallus is endolithic and the paraphyses have distintegrated. This is probably a species of Verrucaria. The lichen was collected by the author and botanist Steven Disparti at Crystal Cove Beach State Park in Orange County, California.
|
|
Crustose Terrestrial Lichens Of North America
A dozen or more species of lichens
may grow on a single boulder, often completely covering the rock
surface. Crustose lichens form such a tight interface with the
crystalline surface of some rocks that they are practically
impossible to scape off. They will even grow on shiny black
obsidian. Many crustose rock lichens spend most of their lives
in a desiccated "dormant" state and have extremely slow annual
growth rates. Lichens dry out very rapidly and may loose up to
98 percent of their water content. When a lichen is wetted by
rain or morning dew, it quickly imbibes water like blotting
paper, and photosynthesis within its little algal partner is
revived for a while. Since lichens are among the first plants to
grow on bare rock, they play a role in soil formation by slowly
etching the rock surface. Microscopic rock fragments intermeshed
with the lichen become loosened by expansion and contraction, as
the lichen is alternately moistened and dried.
On rugged granite outcrops, large
colonies of lime-green map lichen (Rhizocarpon
geographicum), ashy-gray Aspicilia cinerea and orange
Caloplaca saxicola may be thousands of years old. In
fact, the colorful chartreuse rock lichen Acarospora
chlorophana may grow only a few millimeters per century. One
has only to gaze at the spectacular panoramas of glacier-carved
granite throughout the Sierra Nevada to appreciate the magnitude
of some crustose lichens. The blackened surface of massive domes
of Yosemite National Park is actually crustose species of
Buellia, Verrucaria and Lecidea atrobrunnea. Growth rates from rock lichens on glacial moraines have been used to approximate
the time interval between the advance and retreat of glaciers.

|
Left: Spectacular granite domes of Yosemite National Park in California's rugged Sierra Nevada are blackened by colonies of the crustose rock lichen Lecidea atrobrunnea. Close-up view of a hand lens and Lecidea atrobrunnea, a common crustose lichen throughout granite peaks and domes of the Sierra Nevada. The photo was taken at the summit of Polly Dome, the highest dome in Yosemite National Park.
|
Rock lichens have played an
important role in the survival of native people and explorers.
In addition to providing food for their animals, Indians, Eskimos
and Laplanders eat certain lichens. Leafy lichens called rock
tripes (Umbilicaria) are eaten raw and are boiled in soups.
They are also added to salads or deep fried, and are considered a
delicacy in Japan. Throughout history, peasants of Persia have
avoided mass starvation by eating the abundant crustose rock
lichen Lecanora esculenta. This lichen readily becomes
detached in small patches and is blown off the rocks by wind,
often accumulating in crevices and under shrubs. It is mixed
with meal and made into a kind of bread in Turkey and northern
Iran. In fact, some biblical scholars think this lichen may have
been the "manna" which saved the starving Israelites during their
exodus from Egypt.
|
Spectacular colonies of lichens on alpine metamorphic outcrop above Gaylor Lakes in the Sierra Nevada. Some of the lichens include light gray foliose Umbilicaria, dark brown clumps of Acarospora thamnina, bright orange Caloplaca, yellow Candelariella, gray crustose Rhizocarpon disporum, dark brown crustose Staurothele clopima and Lecidea atrobrunnea.
|
Xerarch Succession On Bare Rock
In the natural environment, plants are constantly invading and colonizing new habitats--a phenomenon known as succession. When succession starts on bare rock or ground it is called xerarch succession. Since lichens are among the first plants to colonize bare rock, they play an important role in primary succession. After lichens have etched and crumbled the rock surface for centuries, mineral soil and organic matter begins to accumulate. Then other plants such as mosses and grasses begin to grow, followed by herbs, hardy shrubs, and finally trees. Although lichens produce weak organic (phenolic) acids, it is doubtful that these acids have much effect on the etching of rocks, unless they are calcareous. For most rock surfaces, the etching process is probably mechanical. Crustose rock lichens are able to grow on bare rock, sinking their spreading thallus into every minute nook and cranny. Microscopic rock fragments intermeshed with lichen thallus become loosened by expansion and contraction, as the thallus is alternately moistened and dried. The rock-breaking power of plants is greatly enhanced when seeds fall into cracks and then germinate. This is particularly true in the case of woody shrubs and conifers with powerful expanding root systems. Throughout the subalpine Sierra Nevada there are forests of pine, fir, and hemlock growing in relatively shallow soils and duff overlaying solid granite. This massive bedrock of granite was scoured and polished by glaciers as recently as 12,000 years ago.
Concluding Remarks About Rock Lichens
In addition to desert varnish and lichens, other organisms coat the surfaces of boulders and tree trunks, including cyanobacteria and green algae. Cyanobacteria (formerly called blue-green algae) often form dense, feltlike meshworks of microscopic filaments. They cover the trunks of trees and boulders, particularly in wet regions or shady habitats. In Guatemala, the massive pyramids in the ancient Maya city of Tikal are covered with a thin black layer of cyanobacteria of the genus Scytonema. In forests of southern California, a related species of filamentous cyanobacteria grows on the trunks of young incense cedars. From a distance the trees look as though they were charred by a fire.
|
Left: The limestone outcrop in foreground is covered with a felty layer of dark cyanobacteria (Scytonema). In the distance are Caribbean pines (Pinus caribaea). Photo was taken in the Mountain Pine Ridge area of Belize. Right: Microscopic view (400x) of the cellular filaments of the cyanobacterium Scytonema. The filaments are composed of minute prokaryotic cells within a conspicuous outer sheath. A special cell called a heterocyst can be seen in the upper part of the right filament. In many cyanobacteria this is the site of nitrogen fixation where atmospheric nitrogen is converted into ammonia.
|
Through the remarkable marriage
between algae and fungi, lichens have etched out an existence in
places where no other life forms could possibly live. Although
they have survived millions of years of evolution, many lichen
species are now endangered by atmospheric pollution. This same
fate may also affect some of the desert varnish microbes. Since
lichens absorb most of their mineral nutrients from the air and
rain water, they are especially vulnerable to toxic air-borne
pollutants. Because they have no way to excrete the elements
they absorb into their tissue, toxic compounds become even more
concentrated. The toxins cause the photosynthetic algal cells to
deteriorate and the subsequent death of the fungal spouse.
Extensive off-road vehicle activity in some desert areas stirs up
vast quantities of alkaline dust. This increased atmospheric
alkalinity may affect manganese oxidation by varnish bacteria,
thus slowing or inhibiting the development of desert varnish.
Radioactive fallout is one of the most insidious atmospheric toxins, affecting entire ecosystems through biological magnification. Soil lichens, including reindeer moss (Cladina rangiferina) and Iceland moss (Cetraria islandica), carpet thousands of square miles of North American and Eurasian tundra, providing a major food source for caribou, reindeer and musk ox. Laplanders absorb more radiation than other people on earth because they subsist largely on reindeer, which in turn feed on lichens. In fact, Lapps may concentrate up to one hundred times the level of radioactive isotopes found in tundra lichens. When radioactive clouds from the Chernobyl nuclear power plant meltdown rained cesium 137 on hundreds of miles of reindeer moss, more than 70,000 reindeer had to be destroyed because of dangerous levels of radiation.

|
Deer "moss" (Cladina evansii), a fruticose ground lichen native to sand pine forests (Pinus clausa) of Florida. This genus of intricately-branched fruticose lichens also includes the widespread Arctic reindeer "moss" (C. rangiferina). Several other tundra species of Cladina are also eaten by caribou, reindeer and musk ox, including C. arbuscula, C. mitis, C. stellaris and C. stygia.
|
These same tundra lichens may have played a major role in the migration of people from Siberia to North America across the Bering Land Bridge over 20,000 years ago. If these ancient explorers fed primarily on native animals, which in turn fed on lichens, then the importance of lichens to their remarkable journey is astonishing.
Like the proverbial canary used to
detect invisible but deadly methane fumes in a coal mine, lichens
are sensitive barometers of atmospheric pollution. In fact,
their vulnerability has made them very effective air pollution
monitoring stations for the U.S. Forest Service and National Park
Service. Lichens and desert varnish are also some of the most
fascinating and complex of all living organisms. We still have a
lot to learn from them. In addition, they coat our mountains
with a myriad of beautiful colors.
|
|
References
- Ahmadjian, V. 1967. The Lichen Symbiosis. Blaisdell Publishing Co., Waltham, Massachusetts.
- Armstrong, W.P. 2002. "The Marriage Between Algae and Fungi." Wildflower 18 (1): 10-16.
- Armstrong, W.P. and J.L. Platt 1993. "The Marriage Between Algae and Fungi." Fremontia 22 (2): 3-12.
- Atsatt, P.R. 1988. "Are Vascular Plants Inside Out Lichens?" Ecology: 69 (1): 17-23.
- Atsatt, P.R. 2003. "Fungus Propagules in Plastids: The Mycosome Hypothesis." International Microbiology 6: 17-26.
- Brock, T.M. and M.T. Madigan. 1988. Biology of Microorganisms (5th Edition). Prentice Hall, Englewood Cliffs, New Jersey.
- Brodo, I.M., Sharnoff, S.D. and S. Sharnoff. 2001. Lichens of North America. Yale University Press, New Haven and London.
- Dobson, F. 1979. Lichens: An Illustrated Guide. The Richmond Publishing Co. Ltd., Richmond, Surrey, England.
- Dorn, R.I. 1982. "Enigma of the Desert." Environment Southwest Number 497: 3-5.
- Dorn, R.I. and T.M. Oberlander. 1982. "Rock Varnish." Progress In Physical Geography 6: 317-367.
- Dorn, R.I. and T.M. Oberlander. 1981. "Microbial Origin of Desert Varnish." Science 213: 1245-1247.
- Hale, M.E. and M. Cole. 1988. Lichens of California. University of California Press, Berkeley.
- Hale, M.E. 1969. How to Know the Lichens. Wm. C. Brown Company, Dubuque, Iowa.
- McCune, B. and T.Goward. 1995. Macrolichens of the Northern Rocky Mountains. Mad River Press, Inc., Eureka, California.
- Nash, T.H. III, Ryan, B.D., Gries, C. and F. Bungartz. 2002. Lichen Flora of the Greater Sonoran Desert Region Volume I. Lichens Unlimited, Arizona State University, Tempe, Arizona.
- Nash, T.H. III. 1996. Lichen Biology. Cambridge University Press, Cambridge.
- Purvis, W. 2000. Lichens. Smithsonian Institution Press, Washington, D.C.
- Retallack, G. 1994. "Were the Ediacaran Fossils Lichens?" Paleobiology 20 (4): 523-544.
- Richardson, David H.S. 1974. The Vanishing Lichens. Hafner Press, New York.
- Vitt, D.H., Marsh, J.E. and R.B. Bovey. 1988. Mosses, Lichens & Ferns of Northwest North America. Lone Pine Publishing, Edmonton, Alberta, Canada.
|
|










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}