

Abstract

We compared silver (Ag) bioavailability and toxicity to a freshwater gastropod after exposure to ionic silver (Ag+) and to Ag nanoparticles (Ag NPs) capped with citrate or with humic acid. Silver form, exposure route, and capping agent influence Ag bioaccumulation dynamics in Lymnaea stagnalis. Snails efficiently accumulated Ag from all forms after either aqueous or dietary exposure. For both exposure routes, uptake rates were faster for Ag+ than for Ag NPs. Snails efficiently assimilated Ag from Ag NPs mixed with diatoms (assimilation efficiency (AE) ranged from 49 to 58%) and from diatoms pre-exposed to Ag+ (AE of 73%). In the diet, Ag NPs damaged digestion. Snails ate less and inefficiently processed the ingested food, which adversely impacted their growth. Loss rates of Ag were faster after waterborne exposure to Ag NPs than after exposure to dissolved Ag+. Once Ag was taken up from diet, whether from Ag+ or Ag NPs, Ag was lost extremely slowly. Large Ag body concentrations are thus expected in L. stagnalis after dietborne exposures, especially to citrate-capped Ag NPs. Ingestion of Ag associated with particulate materials appears as the most important vector of uptake. Nanosilver exposure from food might trigger important environmental risks.
Introduction
Methods

 where Bmax (nmol g–1) represents the number of transport sites, [M]exposure is the metal exposure concentration (nmol l–1 or nmol g–1), and Kmetal (nmol l–1 or nmol g–1) represents the affinity of each transport site for a metal.
where Bmax (nmol g–1) represents the number of transport sites, [M]exposure is the metal exposure concentration (nmol l–1 or nmol g–1), and Kmetal (nmol l–1 or nmol g–1) represents the affinity of each transport site for a metal.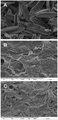
Figure 1. SEM images of (a) diatom mat onto which a low concentration of HA-Ag NPs (17 nmol g–1 of Ag as HA-Ag NPs) was filtered through; (b) feces collected after 48 h of depuration for the lowest (17 nmol g–1) and highest (187 nmol g–1) exposure concentrations to HA-Ag NPs.
Results

Figure 2. Silver uptake rates (nmol g–1 d–1 ± 95% C.I.) in L. stagnalis soft tissue after waterborne and dietborne exposures. Each symbol represents Ag concentrations (from which background Ag was removed) for 10 individuals and 5 water or diatoms samples (±95% C.I.). Open circles are for the dissolved exposure; solid circles are for dietary exposures. Solid lines represent linear regression relationships; dotted lines represent nonlinear regression fits to Michaelis–Menten equation (eq 2). Inset in panels B, E, and F shows data at a smaller scale.
| Ag+ | cit-Ag NPs | HA-Ag NPs | |
|---|---|---|---|
| biodynamic parameters | |||
| kuw (l g–1 d–1)b | 1.1 ± 0.1 (6) | 0.35 ± 0.01 (4) | 1.2 ± 0.2 (3) |
| kuf (g g–1 d–1)b | 0.81 ± 0.03 (6) | 0.10 ± 0.09 (3) | 0.27 ± 0.12 (5) |
| kew (d–1)b | 0.004 ± 0.013 (9) | 0.058 ± 0.019 (9) | 0.051 ± 0.020 (11) |
| kef (d–1)b | 0.005 ± 0.031 (7) | N.D. (7)d | 0.075 ± 0.026 (7) |
| kgw (d–1)c | N.D. (70)d | 0.008 ± 0.005 (92) | 0.015 ± 0.003 (110) |
| kgf (d–1)c | 0.036 ± 0.006 (69) | N.D. (69)d | 0.031 ± 0.006 (78) |
| AE (%)c | 73 ± 5 (44) | 58 ± 8 (27) | 49 ± 7 (30) |
| IR (g g–1 d–1)c | 0.91 ± 0.09 (44) | 0.24 ± 0.07 (27) | 0.29 ± 0.04 (30) |
| defecation rate (g g–1 d–1)c | 0.066 ± 0.005 (64) | 0.041 ± 0.005 (54) | 0.052 ± 0.005 (54) |
| metal binding characteristics | |||
| Bmax (nmol g–1)b | 31 ± 5 (7) | 18 ± 6 (5) | 82 ± 26 (5) |
| Kmetal (nmol l–1)b | 20 ± 7 (7) | 29 ± 23 (5) | 101 ± 54 (5) |
N.D. Not detected.
The number of replicate samples containing 10 individual snails is in parentheses.
The number of individual snails is in parentheses.
Set at 0.001 for modeling.

Figure 3. Proportional loss of Ag over time in L. stagnalis after waterborne and dietborne exposures. Each symbol represents Ag concentrations (from which background Ag was removed) for 10 individuals (±95% C.I.). Open circles are for the dissolved exposure; solid circles are for dietary exposures. Solid lines represent net loss of Ag; dotted lines represent body growth dilution; dashed lines represent the combined loss through elimination and growth dilution.
Discussion
Acknowledgment
Funding for this work was provided from the Toxic Substances Research Program of the U.S. Geological Survey, the National Research Program (USGS) and by a grant from the Leverhulme Trust (F/00 696/N). The authors thank A. D. Dybowska and D. Berhanu for their assistance with particle synthesis and characterization, D. B. Kent for help with the chemical speciation calculations, J. A. Garcia for his assistance with biodynamics experiments as well as A. Kleckner and D. J. Cain for help with algal cultures. Critical comments from D. J. Cain and A. D. Dybowska are greatly appreciated.
Supporting Information
TEM images and size distribution for cit-Ag NPs and HA-Ag NPs, particle characterization, ionic composition of the MOD water, biodynamic model equations, EDTA extraction procedure, SEM images, and pictures of diatom mats onto which Ag NPs were filtered through, metal analysis using ICP-MS, Ag concentrations in L. stagnalis after waterborne and dietborne exposures, Ag AE, food IR, and defecation rates as a function of Ag exposure concentrations, comparative SEM images of feces after dietborne exposures to cit-Ag NPs, and dry weight in L. stagnalis as a function of time during depuration (Figures S1–S8). This material is available free of charge via the Internet at http://pubs.acs.org.
References
This article references 34 other publications.
- 1.The Project on Emerging Nanotechnologies. http:/www.nanotechnology.org/.There is no corresponding record for this reference.
- 2.Mueller
, N. C.; Nowack, B.Exposure modeling of engineered nanoparticles in the environment Environ. Sci. Technol. 2008, 42, 4447– 4453[ACS Full Text ], [CAS]2.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD1cXlslOju7k%253D&md5=9dffbf07c702297356381d2dbaa42e95Mueller, Nicole C.; Nowack, BerndEnvironmental Science & Technology (2008),This work used a life-cycle perspective to model the quantity of engineered nano-particles released into the environment. Three types of nano-particles were studied: nano-Ag, nano-TiO2, and C nanotubes (CNT). Quantification was based on a substance flow anal. from products to air, soil, and water in Switzerland. The following were model inputs: estd. worldwide prodn. vol.; prodn. vol. allocation to product categories; particle release from products; and flow coeffs. within environmental compartments. Predicted environmental concns. (PEC) were then compared to predicted no effect concns. (PNEC) derived from the literature to est. a possible risk. Expected concns. of the 3 nano-particles in different environmental compartments varied widely, due to the different life cycles of the nano-particle-contg. products. PEC values for nano-TiO2 in water were 0.7-16 μg/L, close to or higher than the PNEC value for nano-TiO2 (<1 μg/L). Risk quotients (PEC:PNEC) for CNT and nano-Ag were much <1; thus, comprising no reason to expect adverse effects from those particles. Results made it possible for the first time to perform a quant. environmental risk assessment for nano-particles and suggest further detailed studies of nano-TiO2.
], [CAS]2.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD1cXlslOju7k%253D&md5=9dffbf07c702297356381d2dbaa42e95Mueller, Nicole C.; Nowack, BerndEnvironmental Science & Technology (2008),This work used a life-cycle perspective to model the quantity of engineered nano-particles released into the environment. Three types of nano-particles were studied: nano-Ag, nano-TiO2, and C nanotubes (CNT). Quantification was based on a substance flow anal. from products to air, soil, and water in Switzerland. The following were model inputs: estd. worldwide prodn. vol.; prodn. vol. allocation to product categories; particle release from products; and flow coeffs. within environmental compartments. Predicted environmental concns. (PEC) were then compared to predicted no effect concns. (PNEC) derived from the literature to est. a possible risk. Expected concns. of the 3 nano-particles in different environmental compartments varied widely, due to the different life cycles of the nano-particle-contg. products. PEC values for nano-TiO2 in water were 0.7-16 μg/L, close to or higher than the PNEC value for nano-TiO2 (<1 μg/L). Risk quotients (PEC:PNEC) for CNT and nano-Ag were much <1; thus, comprising no reason to expect adverse effects from those particles. Results made it possible for the first time to perform a quant. environmental risk assessment for nano-particles and suggest further detailed studies of nano-TiO2. - 3.Liau
, S. Y.; Read, D. C.; Pugh, W. J.; Furr, J. R.; Russel, A. D.Interaction of silver nitrate with readily identifiable groups: relationship to the antibacterial action of silver ions Lett. Appl. Microbiol. 1997, 25, 279– 283[CrossRef], [PubMed], [CAS]3.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADyaK2sXntFCktb0%253D&md5=853f4cfa051d67890aa9638f5c2b05bbLiau, S. Y.; Read, D. C.; Pugh, W. J.; Furr, J. R.; Russell, A. D.Letters in Applied Microbiology (1997),Microbiol. it was demonstrated that amino acids, e.g. cysteine (CySH), and other compds., e.g. sodium thioglycollate, which contain thiol groups neutralized the activity of silver nitrate against Pseudomonas aeruginosa PAO1. Amino acids with disulfide bonds were inactive, with the exception of L-cystine di-Me ester, as were all amino acids with no sulfur groups. Iodoacetamide reacted with CySH to produce a CyS-acetamide complex that was unable to quench the activity of Ag+. Chem. analyses using cyclic voltammetry demonstrated that high coordination nos. (3·1) were obtained with thiol-contg. amino acids and low nos. (0·28-0·4) with other amino acids. Both microbiol. and chem., the results imply that interaction of Ag+ with thiol groups plays an essential role in bacterial inactivation. - 4.Ratte
, H. T.Bioaccumulation and toxicity of silver compounds: A review Environ. Toxicol. Chem. 1999, 18, 89– 108[CrossRef], [CAS]4.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADyaK1MXkt1Or&md5=4f96982bc1c71fffb5ceb4fc76bf004eRatte, Hans ToniEnvironmental Toxicology and Chemistry (1999),A review with 123 refs. of the literature revealed that bioaccumulation of silver in soil is rather low, even if the soil is amended with silver-contg. sewage sludge. Plants grown on tailings of silver mines were found to have silver primarily in the root systems. In marine and freshwater systems, the highest reported bioconcn. factors (BCFs) were obsd. in algae (>105), probably because of adsorption of the dissolved silver (<0.45 p.m fraction) to the cell surface. In herbivorous organisms (e.g., zooplankton and bivalves), the BCF was lower by about two orders of magnitude. Low amts. of silver were assimilated from food with no substantial biomagnification. In carnivores (e.g., fish), the BCF was also lower by one order of magnitude with no indication of biomagnification. Toxicity of silver occurs mainly in the aq. phase and depends on the concn. of active, free Ag+ ions. Accordingly, many processes and water characteristics reduce silver toxicity by stopping the formation of free Ag-, binding Ag+, or preventing binding of Ag+ to the reactive surfaces of organisms. The soly. of a silver compd., and the presence of complexing agents (e.g., thiosulfate or chloride), dissolved org. carbon, and competing ions are important. In soil. sewage sludge, and sediments, in which silver sulfide predominates, the toxicity of silver, even at high total concns., is very low. The highly sol. silver thiosulfate complex has low toxicity, which can be attributed to the silver complexed by thiosulfate. Silver nitrate is one of the most toxic silver compds. The toxic potential of silver chloride complexes in seawater is and will be an important issue for investigation. Aquatic chronic tests, long-term tests, and tests including sensitive life stages show lower toxicity thresholds (∼1 μg Ag+/L). The organisms viewed as most sensitive to silver are small aquatic invertebrates, particularly embryonic and larval stages. - 5.Lee
, K. J.; Nallathamby, P. D.; Browning, L. M.; Osgood, C. J.; Xu, X.-H. N.In vivo imaging of transport and biocompatibility of single silver nanoparticles in early development of zebrafish embryos ACS Nano 2007, 1, 133– 143[ACS Full Text], [CAS]5.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD2sXhtFSnsL%252FF&md5=142ed0938263084d43b4631105bbf7c4Lee, Kerry J.; Nallathambly, Prakash D.; Browning, Lauren M.; Osgood, Christopher J.; Xu, Nancy Xiao-HongACS Nano (2007),Real-time study of the transport and biocompatibility of nanomaterials in early embryonic development at single-nanoparticle resoln. can offer new knowledge about the delivery and effects of nanomaterials in vivo and provide new insights into mol. transport mechanisms in developing embryos. In this study, the authors directly characterized the transport of single silver nanoparticles into an in vivo model system (zebrafish embryos) and investigated their effects on early embryonic development at single-nanoparticle resoln. in real time. The authors designed highly purified and stable (not aggregated and no photodecompn.) nanoparticles and developed single-nanoparticle optics and in vivo assays to enable the study. The authors found that single Ag nanoparticles (5-46 nm) are transported into and out of embryos through chorion pore canals (CPGs) and exhibit Brownian diffusion (not active transport), with the diffusion coeff. inside the chorionic space (3 × 10-9 cm2/s) ∼26 times lower than that in egg water (7.7 × 10-8 cm2/s). In contrast, nanoparticles were trapped inside CPCs and the inner mass of the embryos, showing restricted diffusion. Individual Ag nanoparticles were obsd. inside embryos at each developmental stage and in normally developed, deformed, and dead zebrafish, showing that the biocompatibility and toxicity of Ag nanoparticles and types of abnormalities obsd. in zebrafish are highly dependent on the dose of Ag nanoparticles, with a crit. concn. of 0.19 nM. Rates of passive diffusion and accumulation of nanoparticles in embryos are likely responsible for the dose-dependent abnormalities. Unlike other chems., single nanoparticles can be directly imaged inside developing embryos at nanometer spatial resoln., offering new opportunities to unravel the related pathways that lead to the abnormalities. - 6.Luoma
, S. N.; Rainbow, P. S.Why is metal bioaccumulation so variable? Biodynamics as a unifying concept Environ. Sci. Technol. 2005, 39, 1921– 1931[ACS Full Text], [CAS]6.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD2MXhsF2ntLY%253D&md5=1bef4920bafd6639959ca3daab0a3b11Luoma, Samuel N.; Rainbow, Philip S.Environmental Science and Technology (2005),Ecol. risks from metal contaminants are difficult to document because responses differ among species, threats differ among metals, and environmental influences are complex. Unifying concepts are needed to better tie together such complexities. Here we suggest that a biol. based conceptualization, the biodynamic model, provides the necessary unification for a key aspect in risk: metal bioaccumulation (internal exposure). The model is mechanistically based, but empirically considers geochem. influences, biol. differences, and differences among metals. Forecasts from the model agree closely with observations from nature, validating its basic assumptions. The biodynamic metal bioaccumulation model combines targeted, high-quality geochem. analyses from a site of interest with parametrization of key physiol. consts. for a species from that site. The physiol. parameters include metal influx rates from water, influx rates from food, rate consts. of loss, and growth rates (when high). We compiled results from 15 publications that forecast species-specific bioaccumulation, and compare the forecasts to bioaccumulation data from the field. These data consider concns. that cover 7 orders of magnitude. They include 7 metals and 14 species of animals from 3 phyla and 11 marine, estuarine, and freshwater environments. The coeff. of detn. (R2) between forecasts and independently obsd. bioaccumulation from the field was 0.98. Most forecasts agreed with observations within 2-fold. The agreement suggests that the basic assumptions of the biodynamic model are tenable. A unified explanation of metal bioaccumulation sets the stage for a realistic understanding of toxicity and ecol. effects of metals in nature. - 7.Rainbow
, P. S.Trace metal concentrations in aquatic invertebrates: Why and so what? Environ. Pollut. 2002, 120, 497– 507[CrossRef], [PubMed], [CAS]7.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD38XnsF2kt7w%253D&md5=918954067f19d448aff15493081c5b9eRainbow, Philip S.Environmental Pollution (Oxford, United Kingdom) (2002),A review. All aquatic invertebrates take up and accumulate trace metals whether essential or not, and subsequent body concns. of trace metals show enormous variability across metals and invertebrate taxa. Accumulated metal concns. are interpreted in terms of different trace metal accumulation patterns, dividing accumulated metals into two components - metabolically available metal and stored detoxified metal. Crustaceans are used as examples of different accumulation patterns that will have a general applicability to all aquatic invertebrates. Toxicity is related to a threshold concn. of metabolically available metal and not to total accumulated metal concn. The significance of accumulated metal concns. is discussed in terms of the biol. significance, including the attempted recognition of a high or low concn., and of the applied use of aquatic invertebrates in biomonitoring programs assessing geog. and temporal variation in trace metal bioavailabilities in aquatic systems. - 8.Kittler
, S.; Greulich, C.; Diendorf, J.; Köller, M.; Epple, M.Toxicity of silver nanoparticles increases during storage because of slow dissolution under release if silver ions Chem. Mater. 2010, 22, 4548– 4554[ACS Full Text], [CAS]8.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BC3cXpsV2ltbo%253D&md5=4b404823c7060e06919cb666a6318b49Kittler, S.; Greulich, C.; Diendorf, J.; Koeller, M.; Epple, M.Chemistry of Materials (2010),The dissoln. of citrate-stabilized and poly(vinylpyrrolidone)-stabilized silver nanoparticles in water was studied by dialysis for up to 125 days at 5, 25, and 37 °C. The particles slowly dissolve into ions on a time scale of several days. However, in all cases, a limiting value of the released silver was obsd., i.e., the particles did not completely dissolve. In some cases, the nanoparticles released up to 90% of their wt. Formal kinetic data were computed. Rate and degree of dissoln. depended on the functionalization as well as on the storage temp. The release of silver led to a considerably increased toxicity of silver nanoparticles which had been stored in dispersion for several weeks toward human mesenchymal stem cells due to the increased concn. of silver ions. Consequently, "aged" (i.e., immersed) silver nanoparticles are much more toxic to cells than freshly prepd. silver nanoparticles. - 9.Misra
, S. K.; Dybowska, A.; Berhanu, D.; Krisnadasan, S.; Croteau, M.-N.; Luoma, S. N.; Boccaccini, A. R.; Valsami-Jones, E.Surface modification—The key to dissolution and fate of silver nanoparticles. Nano Lett. In preparation.There is no corresponding record for this reference. - 10.Navarro
, E.; Piccapietra, F.; Wagner, B.; Marconi, F.; Kaegi, R.; Odzak, N.; Sigg, L.; Behra, R.Toxicity of silver nanoparticles to Chlamydomonas reinhardtii Environ. Sci. Technol. 2008, 42, 8959– 8964[ACS Full Text], [CAS]10.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD1cXhtFOqt7nO&md5=da32b6e9fb221d6c2943e9388ea374cfNavarro, Enrique; Piccapietra, Flavio; Wagner, Bettina; Marconi, Fabio; Kaegi, Ralf; Odzak, Niksa; Sigg, Laura; Behra, RenataEnvironmental Science & Technology (2008),Silver nanoparticles (AgNP) are likely to enter the aquatic environment because of their multiple uses. The authors have examd. the short-term toxicity of AgNP and ionic silver (Ag+) to photosynthesis in C. reinhardtii using fluorometry. AgNP ranged in size from 10 to 200 nm with most particles around 25 nm. As detd. by DGT (diffusive gradients in thin films), by ion-selective electrode, and by centrifugal ultrafiltration, ∼1% of the AgNP was present as Ag+ ions. Based on total Ag concn., toxicity was 18 times higher for AgNO3 than for AgNP (in terms of EC50). However, when compared as a function of the Ag+ concn., the toxicity of AgNP appeared to be much higher than that of AgNO3. The ionic Ag+ measured in the AgNP suspensions could not fully explain the obsd. toxicity. Cysteine, a strong Ag+ ligand, abolished the inhibitory effects on photosynthesis of both AgNP and Ag+. Together, the results indicate that the interaction of these particles with algae influences the toxicity of AgNP, which is mediated by Ag+. Particles contributed to the toxicity as a source of Ag+ which is formed in the presence of algae. - 11.Garcia-Alonso
, J.; Khan, F. R.; Misra, S. K.; Turmaine, M.; Smith, B. D.; Rainbow, P. S.; Luoma, S. N.; Valsami-Jones, E.Cellular internalization of silver nanoparticles in gut epithelia of the estuarine polychaete Nereis diversicolor. Environ. Sci. Technol. 2011, 45, 4630− 4636.[ACS Full Text], [CAS]11.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BC3MXlt1Sht7s%253D&md5=6614cab3cec327eca913805883a44956Garcia-Alonso, Javier; Khan, Farhan R.; Misra, Superb K.; Turmaine, Mark; Smith, Brian D.; Rainbow, Philip S.; Luoma, Samuel N.; Valsami-Jones, EugeniaEnvironmental Science & Technology (2011),Silver nanoparticles (AgNPs) are widely used which may result in environmental impacts, notably within aquatic ecosystems. As estuarine sediments are sinks for numerous pollutants, but also habitat and food for deposit feeders such as Nereis diversicolor, ingested sediments must be investigated as an important route of uptake for NPs. N. diversicolor were fed sediment spiked with either citrate capped AgNPs (30 ± 5 nm) or aq. Ag for 10 days. Postexposure AgNPs were obsd. in the lumen of exposed animals, and three lines of evidence indicated direct internalization of AgNPs into the gut epithelium. With TEM, electron-dense particles resembling AgNPs were obsd. assocd. with the apical plasma membrane, in endocytic pits and in endosomes. Energy dispersive X-ray anal. (EDX) confirmed the presence of Ag in these particles, which were absent in controls. Subcellular fractionation revealed that Ag accumulated from AgNPs was predominantly assocd. with inorg. granules, organelles, and the heat denatured proteins; whereas dissolved Ag was localized to the metallothionein fraction. Collectively, these results indicate sep. routes of cellular internalization and differing in vivo fates of Ag delivered in dissolved and NP form. For AgNPs an endocytic pathway appears to be a key route of cellular uptake. - 12.Liu
, J; Hurt, R. H.Ion release kinetics and particle persistence in aqueous nano-silver colloids Environ. Sci. Technol. 2010, 44, 2169– 2175[ACS Full Text], [CAS]12.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BC3cXit1Wqsrc%253D&md5=a043ce25387adb8768567c2b35dd76a4Liu, Jingyu; Hurt, Robert H.Environmental Science & Technology (2010),Many important aspects of nanosilver (nAg) are influenced by the ionic activity assocd. with the particle suspension, including antibacterial potency, eukaryotic toxicity, environmental release, and particle persistence. This study synthesizes pure, ion-free, citrate-stabilized nAg colloids as model systems, and measures their time-dependent release of dissolved Ag using centrifugal ultrafiltration and at. absorption spectroscopy. Ion release is shown to be a cooperative oxidn. process requiring both dissolved dioxygen and protons. It produces peroxide intermediates, and proceeds to complete reactive dissoln. under some conditions. Ion release rates increase with temp. (0-37°), and decrease with increasing pH or addn. of humic or fulvic acids. Sea salts have only a minor effect on dissolved Ag release. Ag nanoparticle surfaces can adsorb Ag+, so even simple colloids contain 3 forms of Ag: Ag0 solids, free Ag+ or its complexes, and surface-adsorbed Ag+. Both thermodn. anal. and kinetic measurements indicate that Ag0 nanoparticles will not be persistent in realistic environmental compartments contg. dissolved O. An empirical kinetic law is proposed that reproduces the obsd. effects of dissoln. time, pH, humic/fulvic acid content, and temp. obsd. here in the low range of nAg concn. most relevant for the environment. - 13.Luoma
, S. N.Silver nanotechnologies and the environment: Old problems or new challenges. Project on Emerging Nanotechnologies, Publication 15. Washington DC, 2008.There is no corresponding record for this reference. - 14.Bergsma
, J.; Konings, W. N.The properties of citrate transport in membrane vesicles from Bacillus subtilis Eur. J. Biochem. 1983, 134, 151– 156[CrossRef], [PubMed], [CAS]14.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADyaL3sXkvVersb0%253D&md5=e3fe504547da7a30346857dd49a147e7Bergsma, Jack; Konings, Wil N.European Journal of Biochemistry (1983),The uptake system for citrate is induced in B. subtilis W23 by growth in the presence of citrate and only membrane vesicles isolated from these cells show energy-dependent citrate uptake. Citrate transport in membrane vesicles is strictly dependent on the presence of divalent cations such as Mg2+, Mn2+, Zn2+, Ba2+, Be2+, Ca2+, Cu2+, Co2+, or Ni2+. The initial rate of citrate transport increases with the divalent cation concn. up to a max. The max. initial rate of citrate uptake is reached with 2 mM Mg2+. The cations form stable chelates with citrate. The metal citrate complex is the transported solute. This is demonstrated for citrate uptake in the presence of Ca2+. Membrane vesicles from citrate-grown cells accumulate Ca2+ and citrate only if both solutes are present. Citrate and Ca2+ are accumulated in equimolar quantities. The uptake of Ca2+ but not of citrate is inhibited by Mg2+. Uptake of the metal-citrate complex is inhibited by the uncoupler carbonylcyanide p-trifluoromethoxyphenylhydrazone and in the presence of K+ by valinomycin and nigericin. The inhibitory effects correlate with the effects obsd. on the components of the proton-motive force, indicating that the proton-motive force is a driving force for metal-citrate transport. The no. of protons symported with the metal-citrate complex was detd. under different exptl. conditions from the steady state levels of citrate accumulation, the elec. potential and pH gradient. This no. varies from 1 at pH 4.7 to 2 at pH 8.0. - 15.Chuang
, C.-Y.; Wang, W.-X.Co-transport of metal complexes by the green mussel Perna viridis Environ. Sci. Technol. 2006, 40, 4523– 4527[ACS Full Text], [CAS]15.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD28XlsFaku7k%253D&md5=ce0e46f1756f150d2ccb4690e88b5b95Chuang, Chia-Ying; Wang, Wen-XiongEnvironmental Science & Technology (2006),We examd. the uptake of ligand-bound metals (Cd and Zn) by the green mussel Perna viridis using defined artificial seawater. Different free ion concns. (1 pM to 10 μM) in uptake solns. were created by adding different amts. of total metals (Cd 0.1 nM to 0.1 mM; Zn 0.5 nM to 0.05 mM) and ligands (EDTA, NTA, citric acid). Our results showed that Cd and Zn uptake could not be fully explained by the free Cd and Zn concns. in the presence of different ligands, indicating that metal-ligand complexes were at least partially available for uptake by the mussels. Total Zn concns. appeared to be a better predictor of metal uptake than the free Zn ion concns. in the presence of different ligands. Uptake of lipophilic org. metal complexes was substantially greater than the hydrophilic metal complexes, even though the free ion concn. was comparable or lower. Moreover, the radiolabeled ligand compds. were directly accumulated by the mussels. The accumulation of metal complexes may explain the increased metal uptake with increasing ligand and total metal concn., even though the free ion metal concn. was const. Overall, our exptl. results indicated that free metal ion cannot fully explain metal uptake since metal complex species were also available to the mussels to some extent, apparently through a co-transport process. - 17.Roditi
, H. A.; Fisher, N. S.; Sañudo-Wilhelmy, S. A.Uptake of dissolved organic carbon and trace elements by zebra mussels Nature 2000, 407, 78– 80[CrossRef], [PubMed], [CAS]17.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD3cXmslelur8%253D&md5=d3f33038f18d68f0d7e3c79c93055279Roditi, Hudson A.; Fisher, Nicholas S.; Sanudo-Wilhelmy, Sergio A.Nature (London) (2000),Zebra mussels (Dreissena polymorpha) are widespread and abundant in major freshwater ecosystems in North America, even though the phytoplankton food resources in some of these systems seem to be too low to sustain them. Because phytoplankton biomass is greatly depleted in ecosystems with large D. polymorpha populations and bacteria do not seem to be an important food source for this species, exploitation of alternative carbon sources may explain the unexpected success of D. polymorpha in such environments. Here the authors examine the possibility that absorption of dissolved org. carbon (DOC) from water could provide a nutritional supplement to zebra mussels. The authors find that mussels absorb 14C-labeled DOC produced by cultured diatoms with an efficiency of 0.23%; this indicates that DOC in natural waters could contribute up to 50% of the carbon demand of zebra mussels. The authors also find that zebra mussels absorb some dissolved metals that have been complexed by the DOC; although absorption of dissolved selenium was unaffected by DOC, absorption of dissolved cadmium, silver and mercury by the mussels increased 32-, 8.7- and 3.6-fold, resp., in the presence of high-mol.-wt. DOC. - 18.Voets
, J.; Bervoets, L.; Blust, R.Cadmium bioavailability and accumulation in the presence of humic acid to the Zebra Mussel, Dreissena polymorpha Environ. Sci. Technol. 2004, 38, 1003– 1008[ACS Full Text], [CAS]18.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD2cXjs1WqsQ%253D%253D&md5=316dae8a7024f6b0cfaed8cebcdc6ee1Voets, Judith; Bervoets, Lieven; Blust, RonnyEnvironmental Science and Technology (2004),Metal speciation in aquatic systems is mainly detd. by the type and concn. of ligands present in soln. A very important group of complexing agents is dissolved org. matter (DOM), e.g., humic and fulvic acids. According to the free-ion activity model, only the free metal ion is available to biota. Nevertheless, DOM has been reported to decrease or increase metal uptake, leading to uncertainty concerning the bioavailability of metal-DOM complexes. In this work the effect of Aldrich humic acid on cadmium accumulation by the zebra mussel, Dreissena polymorpha, was studied under lab. conditions. Mussels, collected in a drinking water reservoir, were exposed to varying environmentally relevant concns. of cadmium in the presence and absence of humic acid. Cadmium concns. in the mussel tissues were analyzed, and measurements with a cadmium-ion-selective electrode were made to det. the free cadmium ion activity in the exposure waters. The uptake of humic acid by the zebra mussels was measured by the decrease of the total org. carbon (TOC) concn. in the water over time. The free cadmium ion activity in the water decreased from 51.6% to 19.9% of the total cadmium concn. in the presence of humic acid. This decrease by a factor of 2.6 resulted in a decrease in the cadmium uptake rate in the soft tissue of zebra mussels from 12.9 to 7.9 nmol/g dry wt./day, which corresponds to a decrease by a factor of 1.6. This implies that cadmium uptake rates were higher than predicted by the free-ion activity model and indicates that cadmium-humic acid complexes are partly available to zebra mussels. - 19.Fabrega
, J.; Fawcett, S. R.; Renshaw, J. C.; Lead, J. R.Silver nanoparticle impact on bacterial growth: effect of pH, concentration, and organic matter Environ. Sci. Technol. 2009, 43, 7285– 7290[ACS Full Text], [CAS]19.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD1MXmvFCksrw%253D&md5=7546a2fb7aabc9e90bac890e82c7a070Fabrega, Julia; Fawcett, Shona R.; Renshaw, Joanna C.; Lead, Jamie R.Environmental Science & Technology (2009),Ag nanoparticles (NPs) are widely used as antibacterial agents. This antibacterial property carries with it a potential environmental risk once these NPs are discharged into the environment. This study examd. the impact on Pseudomonas fluorescens over a 24 h exposure of well characterized Ag NPs at pH 6-9, in the presence and absence of Suwannee River humic acids (SRHA). Ag NPs were characterized by size, aggregation, morphol., dissoln., and surface properties under all conditions. Soly. was low (<2%) for all Ag NP concns. (2-2000 ppb) and under all conditions was <40 ppb (0.38μM). SRHA caused a partial disaggregation of Ag NP aggregates by nanoscale film formation, with individual NPs stabilized by charge and entropically driven steric effects. Dissolved Ag reduced bacterial growth entirely at 2000 ppb (19μM) under all conditions and adversely affected growth at 200 ppb (1.9μM) under some conditions, indicating some toxicity. The Ag NPs showed similar toxicity at 2000 ppb (19μM) in the absence of SRHA and at pH 9 only i.e. SRHA mitigated bactericidal action. Soly. and interactions with SRHA indicate that there was a specific nanoparticle effect, which could not be explained by the effect of dissolved Ag. - 20.Parkhurst
, D. L.; Appelo, C. A. J.User’s guide to PHREEQC (version 2) – A computer program for speciation, batch-reaction, one-dimensional transport, and inverse geochemical calculations. Water Resour. Invest. Rept., 99–4259, Northborough, MA. 1999.There is no corresponding record for this reference. - 21.Wen
, L.-S.; Santschi, P. H.; Gill, G. A.; Tang, D.Silver concentrations in Colorado, USA, watersheds using improved methodology Environ. Toxicol. Chem. 2002, 21, 2040– 2051[CrossRef], [PubMed], [CAS]21.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD38XntlGjt7o%253D&md5=5074f6ea2cf056c54b77139980dd6913Wen, Liang-Saw; Santschi, Peter H.; Gill, Gary A.; Tang, DeguiEnvironmental Toxicology and Chemistry (2002),River water samples were collected at 5 sites in the state of Colorado to assess the impact of municipal and industrial discharges on Ag concns. and speciation in surface waters. Samples were collected and analyzed for total (unfiltered collections), filtered (0.1 and 0.4 μm), particulate (≥0.45 μm), and colloidal Ag (3 KDa-0.1 μm) using ultraclean protocols. A series of lab. expts. were conducted to assess bias from sample storage, digestion, and preconcn. protocols. In general, upstream unfiltered and particulate Ag concns. fell within a fairly narrow range, 3.1-21 ng/L and 0.2-1.7 μg/g, resp. Downstream unfiltered and particulate Ag concns. showed a more broad range, 2.8-1110 ng/L and 0.5-104 μg/g, resp., and reflected attenuated impacts of Ag-laden discharge effluents. However, Ag concns. in the 0.1-μm filter-passing fraction 0.8-1.2 Km downstream from major treatment plant effluents were all below the chronic Ag criteria. On av., >60% of the 0.1-μm filter-passing Ag was assocd. with colloidal macromol. org. matter. Ag concns. in colloids (μg/g) were, on av., the same as those in suspended particulate matter. The percentage abundance of colloidal Ag was similar to that of dissolved org. C, suggesting that strong Ag binding ligands exist in both the colloidal and the particle size fractions, as these macromol. ligands likely play a major role in Ag speciation. - 22.Irving
, E. C.; Baird, D. J.; Culp, J. M.Ecotoxicological responses of the mayfly Baetis tricaudatus to dietary and waterborne cadmium: Implications for toxicity testing Environ. Toxicol. Chem. 2003, 22, 1058– 1064[CrossRef], [PubMed], [CAS]22.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD3sXjtFCmsro%253D&md5=e90afd6698d4a5c6727138c9b16412ceIrving, Elaine C.; Baird, Donald J.; Culp, Joseph M.Environmental Toxicology and Chemistry (2003),Trace metals readily accumulated by stream periphyton may enter aquatic food chains through grazer ingestion. Hence, expts. were conducted to det. the ecotoxicol. responses of the grazing mayfly B. tricaudatus to dietary cadmium. Short-term feeding expts. indicated that B. tricaudatus nymphs did not initially avoid grazing on cadmium-contaminated diatom mats. During a partial life-cycle expt., 4 and 10 μg/g of dietary cadmium significantly inhibited grazing whereas 10 μg/g significantly inhibited growth. Feeding inhibition was the likely mechanism that inhibited growth (i.e., through reduced energy intake). Conversely, when exposed to waterborne cadmium using lethal toxicity test procedures, B. tricaudatus nymphs were relatively tolerant (96-h median lethal concn., 1.611 μg/L). Thus, sublethal responses to dietary exposure appeared to be more sensitive than lethal responses to waterborne exposure. Because adult mayfly fecundity is a function of nymph size at emergence, dietary cadmium exposure could increase the extinction probability within mayfly populations. The present study highlights the importance of dietary exposure routes in detg. the ecotoxicol. responses of an organism to a contaminant. Furthermore, the findings emphasize the advantage of evaluating a combination of ecol. relevant, lethal, and sublethal endpoints in lab. methods used to generate data for ecol. risk assessment and regulation. - 23.U.S. Environmental Protection Agency: Methods for Measuring the Acute Toxicity of Effluents and Receiving Waters to Freshwater and Marine Organisms; U.S. EPA: Washington DC, 2002; EPA-821-R-02–012.There is no corresponding record for this reference.
- 24.Croteau
, M.-N.; Dybowska, A. D.; Luoma, S. N.; Valsami-Jones, E.A novel approach reveals that zinc oxide nanoparticles are bioavailable and toxic after dietary exposures Nanotoxicology 2011, 5, 79– 90[CrossRef], [PubMed], [CAS]24.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BC3MXhtVKqsrvI&md5=7ff0c904963df85fe97616293e143342Croteau, Marie-Noele; Dybowska, Agnieszka D.; Luoma, Samuel N.; Valsami-Jones, EugeniaNanotoxicology (2011),If engineered nanomaterials are released into the environment, some are likely to end up assocd. with the food of animals due to aggregation and sorption processes. However, few studies have considered dietary exposure of nanomaterials. Here we show that zinc (Zn) from isotopically modified 67ZnO particles is efficiently assimilated by freshwater snails when ingested with food. The 67Zn from nano-sized 67ZnO appears as bioavailable as 67Zn internalized by diatoms. Apparent agglomeration of the zinc oxide (ZnO) particles did not reduce bioavailability, nor preclude toxicity. In the diet, ZnO nanoparticles damage digestion: snails ate less, defecated less and inefficiently processed the ingested food when exposed to high concns. of ZnO. It was not clear whether the toxicity was due to the high Zn dose achieved with nanoparticles or to the ZnO nanoparticles themselves. Further study of exposure from nanoparticles in food would greatly benefit assessment of ecol. and human health risks. - 25.Croteau
, M.-N.; Luoma, S. N.; Pellet, B.Determining metal assimilation efficiency in aquatic invertebrates using enriched stable metal isotope tracers Aquat. Toxicol. 2007, 83, 116– 125[CrossRef], [PubMed], [CAS]25.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD2sXlsFKjt70%253D&md5=4cb30ecd3bc1830500eac7c6a0e92c6eCroteau, Marie-Noele; Luoma, Samuel N.; Pellet, BastienAquatic Toxicology (2007),We employ a novel approach that combines pulse-chase feeding and multi-labeled stable isotopes to det. gut passage time (GPT), gut retention time (GRT), food ingestion rate (IR) and assimilation efficiency (AE) of three trace elements for a freshwater gastropod. Lettuce isotopically enriched in 53Cr, 65Cu and 106Cd was fed for 2 h to Lymnaea stagnalis. The release of tracers in feces and water was monitored for 48 h, during which unlabeled lettuce was provided ad libidum. The first defecation of 53Cr occurred after 5 h of depuration (GPT), whereas 90% of the ingested 53Cr was recovered in the feces after 22.5 h of depuration (GRT). 53Chromium was not significantly accumulated in the soft tissues upon exposure. In contrast, 65Cu and 106Cd assimilation was detectable for most exptl. snails, i.e., 65/63Cu and 106/114Cd ratios in exposed snails were higher than those for controls. Food IR during the labeled feeding phase was 0.16±0.07 g g-1 d-1. IR was inferred from the amt. of 53Cr egested in the feces during depuration and the concn. of 53Cr in the labeled lettuce. Assimilation efficiencies (±95% CI) detd. using mass balance calcns. were 84±4% for Cu and 85±3% for Cd. The ratio method yields similar AE ests. Expanding the application of this novel stable isotope tracer technique to other metals in a wide variety of species will provide unique opportunities to evaluate the interplay between digestive processes and dietary influx of metals. Understanding the biol. processes that modulate diet-borne metal uptake is crucial to assess the toxicity of diet-borne metals. - 26.Adams
, N. W. H.; Kramer, J. R.Silver speciation in wastewater effluent, surface waters, and pore waters Environ. Toxicol. Chem. 1999, 18, 2667– 2673[CrossRef], [CAS]26.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADyaK1MXns1GlsL8%253D&md5=c23e74b48931c45d8644328d2f200a4dAdams, Nicholas W. H.; Kramer, James R.Environmental Toxicology and Chemistry (1999),Ag, inorg. sulfide, and thiol compds. were measured in municipal wastewater effluent, receiving waters, and pore waters from an anoxic lake sediment in order to predict Ag speciation in these systems. We found submicromolar concns. of inorg. sulfide even in fully oxic surface water. This inorg. sulfide is likely to exist in the form of colloidal metal sulfides, which have been shown to be stable under oxidizing conditions for periods of several hours. Inorg. sulfide in both the wastewater effluent and receiving waters was 200-300 times in excess of Ag concns., whereas inorg. sulfide in the pore waters was 1000-15,000 times in excess of Ag concns. With sulfide in excess of Ag, we predict Ag sulfide complexes to dominate Ag speciation. Thiols were present at low nanomolar levels in pore waters but were not detectable (<1nM) in wastewater effluent or receiving waters. Thiols do not appear to be important to Ag speciation in these freshwater systems. Partitioning of Ag into particulate (>0.45 μm), colloidal (10 kDa, 0.45 μm), and dissolved (<10 kDa) size fractions showed that a significant proportion of Ag is in the colloidal (30-35%) and dissolved phases (15-20%). Dissolved phase concns. were relatively const. in the treatment plant effluent and receiving waters, suggesting that Ag in the <10-kDa size fraction is strongly complexed by ligands that are not significantly affected by aggregation or sorption processes. - 27.Gratton
, S. E.; Ropp, P. A.; Pohlhaus, P. D.; Luft, J. C.; Madden, V. J.; Napier, M. E.; DeSimone, J. M.The effect of particle design on cellular internalization pathways Proc. Natl. Acad. Sci. 2008, 105, 11613– 11618[CrossRef], [PubMed], [CAS]27.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD1cXhtVCnur%252FI&md5=027a7b45c12f0c2fc6364a57f4ba425eGratton, Stephanie E. A.; Ropp, Patricia A.; Pohlhaus, Patrick D.; Luft, J. Christopher; Madden, Victoria J.; Napier, Mary E.; De Simone, Joseph M.Proceedings of the National Academy of Sciences of the United States of America (2008),The interaction of particles with cells is known to be strongly influenced by particle size, but little is known about the interde- pendent role that size, shape, and surface chem. have on cellular internalization and intracellular trafficking. We report on the internalization of specially designed, monodisperse hydrogel particles into HeLa cells as a function of size, shape, and surface charge. We employ a top-down particle fabrication technique called PRINT that is able to generate uniform populations of org. micro- and nanoparticles with complete control of size, shape, and surface chem. Evidence of particle internalization was obtained by using conventional biol. techniques and transmission electron microscopy. These findings suggest that HeLa cells readily internalize nonspherical particles with dimensions as large as 3 μm by using several different mechanisms of endocytosis. Moreover, it was found that rod-like particles enjoy an appreciable advantage when it comes to internalization rates, reminiscent of the advantage that many rod-like bacteria have for internalization in nonphagocytic cells. - 28.Chithrani
, B. D.; Ghazani, A. A.; Chan, W. C. W.Determining the size and shape dependence of gold nanoparticle uptake into mammalian cells Nano Lett. 2006, 6, 662– 668[ACS Full Text], [CAS]28.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD28XhvVCjsLk%253D&md5=8238db28c10da7b816d5fd3f45a16d14Chithrani, B. Devika; Ghazani, Arezou A.; Chan, Warren C. W.Nano Letters (2006),We investigated the intracellular uptake of different sized and shaped colloidal gold nanoparticles. We showed that kinetics and satn. concns. are highly dependent upon the phys. dimensions of the nanoparticles (e.g., uptake half-life of 14, 50, and 74 nm nanoparticles is 2.10, 1.90, and 2.24 h, resp.). The findings from this study will have implications in the chem. design of nanostructures for biomedical applications (e.g., tuning intracellular delivery rates and amts. by nanoscale dimensions and engineering complex, multifunctional nanostructures for imaging and therapeutics). - 29.Cheung
, M.; Wang, W.-X.Influence of subcellular metal compartmentalization in different prey on the transfer of metals to a predatory gastropod Mar. Ecol.: Prog. Ser. 2005, 286, 155– 166[CrossRef], [CAS]29.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD2MXjsVyitrs%253D&md5=dfddaebdd14eabe400b29c74eeae7174Cheung, Mashan; Wang, Wen-XiongMarine Ecology: Progress Series (2005),Marine snails Thais clavigera accumulate high metal concns. in their bodies and trophic transfer may play an important role in metal bioaccumulation in this top predator in the intertidal rocky shore. In this study, the authors examd. the relationship between the subcellular metal distribution in prey (barnacles, mussels, oysters, herbivorous snails, limpets) and the assimilation efficiency (AE) of cadmium, silver and zinc in T. clavigera. The metal subcellular distributions in the oyster prey were also modified and the metal AEs in T. clavigera measured. The authors further purified the subcellular fractions of different prey and detd. how the biochem. properties of different fractions affected the metal AEs. Results showed that the AEs of each metal from different prey were comparable and generally high (50-90%). Significant pos. correlation was found between the Cd AEs and the metallothionein-like protein (MTLP) fraction, and between the AEs of Zn and the trophically available metal (TAM) fraction. For Ag, no relationship between its AE and any of the subcellular fractions was documented. When T. clavigera fed on oysters with different exposure regimes, the metal AEs were not significantly correlated with the metal subcellular partitioning in the oysters. Furthermore, the AEs of Ag, Cd and Zn were similar among the different biochem. fractions. For these 3 metals, T. clavigera assimilated 60-85% from the insol. fraction, 40-80% from the heat-sensitive protein fraction, 45-90% from the MTLP fraction, and 40-80% from the metal-rich granule (MRG) fraction. The results imply that metals bound to the insol. fraction or MRG were also bioavailable to T. clavigera. This study may help to better understand the mechanisms of metal transfer along the intertidal food chain. - 30.Wang
, W.-X.; Fisher, N. S.; Luoma, S. N.Assimilation of trace elements ingested by the mussel Mytilus edulis: Effects of algal food abundance Mar. Ecol.: Prog. Ser. 1995, 129, 165– 176[CrossRef], [CAS]30.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADyaK28Xhs1ejtLk%253D&md5=8e5c1da06bc3aa019e92aed75d1fa9e0Wang, Wen-Xiong; Fisher, Nicholas S.; Luoma, Samuel N.Marine Ecology: Progress Series (1995),Pulse-chase feeding and multi-labeled radiotracer techniques were employed to measure the assimilation of 6 trace elements (110mAg, 241Am, 109Cd, 57Co, 75Se and 65Zn) from ingested diatoms in the mussel Mytilus edulis feeding at different rates (0.1, 0.49 and 1.5 mg dry wt/h). Uniformly radiolabeled diatoms Thalassiosira pseudonana were fed to mussels for 0.5 h, and the behavior of the radiotracers in individual mussels was followed for 96 h in a depuration seawater system. Assimilation efficiency (AE) of each element declined with increasing ingestion rate and increased with gut passage time. The importance of extracellular digestion relative to intracellular digestion increased with ingestion activity, which, when coupled with a decline in AE, suggested that extracellular digestion is less efficient in metal absorption. Zn assimilation was most affected by ingestion rate, suggesting that AE may play a role in the physiol. regulation of this metal in M. edulis. In an expt. to simulate the effects of an acidic gut, lowered pH (5.5) enhanced the release of elements from intact diatom cells, esp. at low particle concn. These results indicate that both feeding components of the mussel (i.e. gut passage time, digestive partitioning) and metal chem. (i.e. metal release at lowered pH within the bivalve gut) are responsible for the difference in the assimilation of trace metals at different food quantities obsd. in mussels. - 31.Reinfelder
, J. R.; Fisher, N. S.The assimilation of elements ingested by marine copepods Science 1991, 251, 794– 796[CrossRef], [PubMed], [CAS]31.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADyaK3MXhtFyrtbg%253D&md5=878cec6c7220f970ffec2b8f60605224Reinfelder, John R.; Fisher, Nicholas S.Science (Washington, DC, United States) (1991),The efficiency with which a variety of ingested elements (Ag, Am, C, Cd, P, S, Se, and Zn) were assimilated in marine calanoid copepods fed uniformly radiolabeled diatoms ranged from 0.9% for Am to 97.1% for Se. Assimilation efficiencies were directly related to the cytoplasmic content of the diatoms. This relation indicates that the animals obtained nearly all their nutrition from this source. The results suggest that these zooplankton, which have short gut resisence times, have developed a gut lining and digestive strategy that provides for assimilation of only sol. material. Because of fraction of total cellular protein in the cytoplasm of the diatoms increased markedly with culture age, copepods feeding on senescent cells should obtain more protein than those feeding on rapidly dividing cells. Elements that are appreciably incorporated into algal cytoplasm and assimilated in zooplankton should be recycled in surface waters and have longer oceanic residence times than elements bound to cell surfaces. - 32.Shoults-Wilson
, W. A.; Zhurbich, O. I.; McNear, D. H.; Tsyusko, O. V.; Bertsch, P.; Unrine, J. M.Evidence for avoidance of Ag nanoparticles by earthworms (Eisenia fetida) Ecotoxicology 2011, 20, 385– 396[CrossRef], [PubMed], [CAS]32.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BC3MXhvVWqurY%253D&md5=a461e8d4816df08924e5db47a6f72918Shoults-Wilson, W. A.; Zhurbich, Oksana I.; McNear, David H.; Tsyusko, Olga V.; Bertsch, Paul M.; Unrine, Jason M.Ecotoxicology (2011),Silver nanoparticles have been incorporated into a wide variety of consumer products, ideally acting as antimicrobial agents. Silver exposure has long been known to cause toxic effects to a wide variety of organisms, making large scale prodn. of silver nanoparticles a potential hazard to environmental systems. Here we describe the first evidence that an organism may be able to sense manufd. nanoparticles in a complex, environmentally relevant exposure and that the presence of nanoparticles alters the organism's behavior. We found that earthworms (Eisenia fetida) consistently avoid soils contg. silver nanoparticles and AgNO3 at similar concns. of Ag. However, avoidance of silver nanoparticles occurred over 48 h, while avoidance of AgNO3 was immediate. It was detd. that avoidance of silver nanoparticles could not be explained by release of silver ions or any changes in microbial communities caused by the introduction of Ag. This leads us to conclude that the earthworms were in some way sensing the presence of nanoparticles over the course of a 48 h exposure and choosing to avoid exposure to them. Our results demonstrate that nanoparticle interactions with organisms may be unpredictable and that these interactions may result in ecol. significant effects on behavior at environmentally relevant concns. - 33.Croteau
, M.-N.; Luoma, S. N.Predicting dietborne metal toxicity from metal influxes Environ. Sci. Technol. 2009, 43, 4915– 4921[ACS Full Text], [CAS]33.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADC%252BD1MXms1Kmsbg%253D&md5=05cd322a06a649faef89be11877bd68bCroteau, Marie-Noele; Luoma, Samuel N.Environmental Science & Technology (2009),Dietborne metal uptake prevails for many species in nature. However, the links between dietary metal exposure and toxicity are not well understood. Sources of uncertainty include the lack of suitable tracers to quantify exposure for metals such as copper, the difficulty to assess dietary processes such as food ingestion rate, and the complexity to link metal bioaccumulation and effects. The authors characterized dietborne copper, nickel, and cadmium influxes in a freshwater gastropod exposed to diatoms labeled with enriched stable metal isotopes. Metal influxes in Lymnaea stagnalis correlated linearly with dietborne metal concns. over a range encompassing most environmental exposures. Dietary Cd and Ni uptake rate consts. (kuf) were, resp., 3.3 and 2.3 times higher than that for Cu. Detoxification rate consts. (kdetox) were similar among metals and appeared 100 times higher than efflux rate consts. (ke). Extremely high Cu concns. reduced feeding rates, causing the relationship between exposure and influx to deviate from linearity; i.e., Cu uptake rates leveled off between 1500 and 1800 nmol g-1 day-1. L. stagnalis rapidly takes up Cu, Cd, and Ni from food but detoxifies the accumulated metals, instead of reducing uptake or intensifying excretion. Above a threshold uptake rate, however, the detoxification capabilities of L. stagnalis are overwhelmed. - 34.Decho
, A. W.; Luoma, S. N.Flexible digestion strategies and trace metal assimilation in marine bivalves Limnol. Oceanogr. 1996, 41, 568– 572[CrossRef], [CAS]34.https://chemport.cas.org/services/resolver?origin=ACS&resolution=options&coi=1%3ACAS%3A528%3ADyaK28XkvFKhtL8%253D&md5=044bbb0680a1839bc142f906342ccd06Decho, Alan W.; Luoma, Samuel N.Limnology and Oceanography (1996),Pulse-chase expts. show that two marine bivalves take optimal advantage of different types of particulate food by varying food retention time in a flexible two-phase digestive system. For example, carbon is efficiently assimilated from bacteria by subjecting nearly all the ingested bacteria to prolonged digestion. Prolonging digestion also enhances assimilation of metals, many of which are toxic in minute quantities if they are biol. available. Detritus-feeding aquatic organisms have always lived in environments naturally rich in particle-reactive metals. We suggest that avoiding excess assimilation of metals could be a factor in the evolution of digestion strategies. We tested that suggestion by studying digestion of particles contg. different Cr concns. We show that bivalves are capable of modifying the digestive processing of food to reduce exposure to high, biol. available, Cr concns. The evolution of a mechanism in some species to avoid high concns. of metals in food could influence how effects of modern metal pollution are manifested in marine ecosystems.













