Nanotechnology Center for Mechanics in Regenerative Medicine2007 Progress Report – Executive SummaryTheme: Analyze and Control Nanoscale Force & Geometry Sensing and Responses in Health and Disease Developments in advanced nanotechnologies have opened new opportunities for the diagnosis and treatment of disease at several different levels. In the first stage that is currently underway, micro- and nanofabricated materials are finding direct application in diagnosing, imaging and treating disease. In a second phase that is the subject of this grant, nanotechnologies provide totally new avenues to systematically analyze the physical aspects of biological nanosystems and cellular functions, enabling one to define nanometer (molecular) level mechanical functions and to then integrate to understand higher level cellular phenomena. A deeper understanding of the engineering principles of biological systems on the nanometer scale and how physical factors co-regulate cellular activities is likely to result in major progress toward understanding how cells function within the complex organism. In a third, later phase, it is expected that this understanding of how cells function in health and disease will open unexpected doors for major innovations in medical diagnosis, drug development, the treatment of disease and regenerative medicine. Although some organs may be too complex to be engineered in vitro, there may be new ways to augment the regenerative processes if we have a better understanding of the normal cellular mechanisms of motility and repair. Indeed many of the promising strategies for treatment of diseases involve stimulation of stem cell expansion, recruitment and differentiation to repair a damaged region of a tissue. Growth, migration and differentiation of the stem cells can be affected by hormone activation but there is an increasing body of evidence showing that matrix mechanical factors play a major role in all three processes. For example, just changing the rigidity of the matrix can cause stem cells to differentiate into totally different tissues (Discher Lab, In Press) and similar changes in breast cancer cell behavior have been noted. Thus, it is very important to understand how mechanical factors are sensed by cells and how they are processed over time. Diseases involved with mechanical disfunction include cancer, cardiac hypertrophy, genetic malformations and immune disorders.
Figure 1: Cellular processes of mechanosensing and responses. This diagram shows the steps in mechanosensing over time that involve periodic testing of the substrate, substrate modification and changes in cellular protein content. Initially, cells will sense the mechanical features of their environment, which will cause rapid motility and signalling responses. As the cell pulls, it will modify the extracellular matrix creating new signals, such as those originating from fibronectin unfolding. Intracellular signals will alter the expression pattern of the cell and, over time, the cellular forces and cellularly generated matrices will change the cell shape. At any stage, extracellular signals, such as hormones or external mechanical stimuli, can cause acute changes that will set off a further round of cell and matrix modifications. From Vogel and Sheetz, 2006 Nat Rev CMB 7:265. We have suggested that the mechanical factors primarily sensed by cells are force and geometry at the molecular level (Vogel and Sheetz, 2006) and changes in those physical factors are transduced into biochemical signals that then activate cellular responses both for fibroblasts and immune cells. Many studies from our labs and others have shown that cells sense forces very rapidly (seconds or less) and the earliest responses are also on a second timescale. Those early force-sensing events appear to involve cytoskeletal protein stretch both externally (Vogel, 2006) and internally (Tamada et al., 2004 Dev. Cell). Internal responses to cell stretch involve downstream tyrosine phosphorylation as a common signaling mechanism in force responsiveness. In responding to forces, cells often generate counter forces that start another cycle of sensing and response. Thus, after a day in culture, a cell may undergo hundreds of sensing and response cycles and may differentiate to become a totally different cell after that period. To understand how to control cell behavior through the mechanical environment, will require a much deeper understanding of the processes of mechanosensing and response. The task of understanding cellular motile functions seems nearly impossible in the light of the large number of proteins involved and the large number of their post-translational modifications. However, quantitative analyses of motility have revealed a relatively few number of distinct motile behaviors and the transitions between these behaviors can be quite abrupt. We postulate that there may be only 10-20 distinct motile functions and each of those could be activated in a variety of cell types. Different motility in different cells is then the result of the different control mechanisms that activate different motile states. This is consistent with the way that many macroscopic machines are engineered; ie, many of the same functional modules are used but overall function is dramatically different because of differences in the numbers of modules and their activation mechanisms. The characterization of each type of motility is potentially very important because it could be critical for normal healing or for causing a disease.
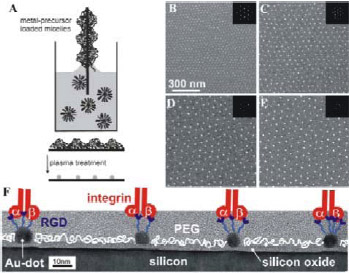
Figure 2: Micellar block copolymer lithography and biofunctionalization. (A) Scheme of diblock copolymer micelle lithography. (B-E) Extended Au-nanodot pattern are displayed by scanning electron microscopy.[43] Uniform Au-nanodots (bright spots) of (B) 3 nm by PS(190)-b- P[2VP(HAuCl4)0.5](190), (C) 5 nm by PS(500)-b- P[2VP(HAuCl4)0.5](270), (D) 6 nm by PS(990)-b- P[2VP(HAuCl4)0.5](385), and (E) 8 nm by PS(1350)-b- P[2VP(HAuCl4)0.5](400) deposited onto Si-wafers are shown. The Au-dots are presented as side view micrographs of a high resolution transmission electron microscope. At the organizational level, the Center for Mechanical Biology is linked to the Motility Consortium (headed by Rick Horwitz) and the other Nanomedicine Centers through a number of important scientific collaborations from fabrication of nanometer-scale materials to a database on the protein interactions in adhesion sites. Sharing of reagents has been an important component for many studies and the open environment enables any ideas to quickly be tested. Our External Advisory Board of Drs. Shu Chien, Barry Coller, and Robert Langer will help us to target the most important applications of the new technologies that emerge from these studies. Previous studies have focused on the early steps in the processes of mechanical sensing and response, using macro- or at best micro-engineered materials. With nanopatterned surfaces, we are now able to systematically test the effect of ligand spacing on cell function (Spatz and Geiger, see Figure 2 for images of hexagonal arrays and other ordered arrays are being fabricated by e-beam lithography by Drs Wind and Hone). Effects of fiber curvature and surface morphology can also be systematically analyzed with current fabrication techniques. Further, the development of active surfaces that can generate force enable us to test for effects of changes in force in subcellular regions. We have plans for adapting of these materials to the study of motility in 3-D. Nanoimprint lithography will enable us to make many copies of surface arrays for multiple cell experiments and to provide these surfaces for other labs to analyze the effects of these surfaces on their cells of interest. Thus, we have the capability now to produce cellular supports with a wide range of nanopatterned morphologies that can be analyzed at the light microscope level. The longer term effects of these materials on cellular adhesion sites, extracellular matrix assembly and cell force generation can be measured in many contexts, including metastatic cell motility, immune synapse formation and stem cell differentiation. The major question in the short term studies is the basic mechanism of mechanosensing and response. In the longer term studies, the integration of the short-term responses overtime will be the focus. Quantitative image processing tools developed in the Wiggins lab will be important for those studies because we will want to quantify the dynamics of the adhesion site components. The lifetimes of the complexes measured by those techniques will be important for modeling the dynamic changes of the adhesion components (Iyengar Lab) and the biochemical changes that might underlie the process of integration. With such quantitative models to predict the overall behavior, we can make specific modifications of components to test the predictions. An important component of the mechanical environment of the cell is the extracellular matrix and we have developed new tools for measuring the upper limits of forces that can be developed or sustained by cells in 2-D and 3-D fiber systems. Magnetic beads are capable of applying nanonewtons of force to fibers and can be used to calibrate the force-dependence of the changes in FRET that accompany the unfolding of fibronectin molecules in the fibers. In addition, we are fabricating new devices to support engineered fibers in organized arrays that can mimic basement membrane and other tissue matrices. The preliminary studies show that the unfolding of the matrix fibers is an important component in the cellular response to the fibers. New active surfaces will enable us to measure the time course of the cellular responses to unfolded matrices and to rapidly change the degree of unfolding. Many of the techniques that are being applied to cells on 2-D matrices will be applied to these new matrices to define the molecular bases of the motile functions that are controlled by matrix.
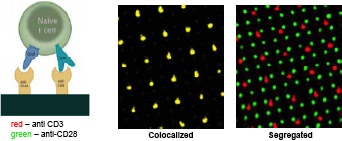
Figure 3: Colocalized surfaces contained 2 μm dots, spaced on 7 um apart on a square lattice, comprised of antibodies to both CD3 and CD28. For segregated surfaces, the anti-CD28 patterns consisted of 1 μm dots on a 3.5 μm grid. Many recent studies have shown that the immune synapse is formed by active movement of liganded receptors through the membrane and it seems that physical separation of components is critical for proper processing of the displayed antigens to give a normal cellular response. Many of the tools that we are developing can be used to address important questions related to the mechanisms by which T-cells probe adjacent membranes. One particular question is whether controlling the distance between components on the micrometer to nanometer level will affect their interactions. As seen in Figure 3, we can now pattern different molecules on the same surface either at separate locations or overlapping. This will tell us whether the receptors need to be interacting for downstream processing events to occur. Such an understanding of the steps in the process will make it possible to target specific aspects of function in autoimmune or transplant situations. To define the protein modules involved in these functions, we are using SiRNA screens and bioinformatics analyses with subsequent experimental testing. From the screens, we will know which proteins are involved. The databases will enable us to rapidly target other potential components of the modules and experiments will clarify the actual changes in function that accompany the loss of given modules. Based upon that understanding we can develop new strategies and reagents for promoting the restoration of proper functions in cells and tissues or for inhibiting function in cases where it has gone awry. The collaborative process that underlies the work of this center has been exciting for many of the center members. We can move rapidly from hypotheses to critical experiments with new tools and reagents that would have taken months to years to generate independently. All of the preliminary studies indicate that the mechanical factors are critical for proper cell function and we are rapidly approaching a level where we can target specific force sensing pathways to perhaps alter cardiac hypertrophy or motility to alter immune functions. |
This page last reviewed: June 25, 2008
Division of Program Coordination, Planning, and Strategic Initiatives • National Institutes of Health • Bethesda, Maryland 20892
![]()
![]()
![]()