|
| |
||
| |
|||||||||||||||||
|
|||||||||||||||||
|
|||||||||||||||||
|
EID Home | Ahead of Print | Past Issues | EID Search | Contact Us | Announcements | Suggested Citation | Submit Manuscript
|
|
Dispatch Sequencing and Staphylococci IdentificationAlexander Mellmann,* Suggested citation for this article
The emerging clinical importance of Staphylococcus aureus and coagulase-negative staphylococci (1) in connection with the expanding number of staphylococcal subspecies described requires accurate identification to the subspecies level. Currently, the genus Staphylococcus is divided into 36 species and 21 subspecies. Staphylococcal subspecies not included in the databases of commercial identification systems, as well as phenotypic variants (e.g., small-colony variants), are often misidentified (2). We recently described the usefulness of genotypic identification of staphylococcal subspecies by using partial 16S rDNA sequences in comparison with phenotypic tests (3). However, the partial 16S rDNA sequences used were not discriminative enough to differentiate all staphylococcal subspecies. When searching for a molecular target for discrimination of staphylococci, several genes have been evaluated, e.g., heat shock protein 60 (hsp60) (4), superoxide dismutase A (sodA) (5), and RNA polymerase B (rpoB) (6). However, these studies concentrated only on a limited number of staphylococcal species. Therefore, a complete reference database of partial rpoB gene sequences from type strains (n = 47) and other culture collection strains, including all validly described staphylococcal subspecies, was created for this study. This reference database was then evaluated with clinical isolates. Results were compared with those previously obtained by 16S rDNA sequencing and conventional phenotypic tests. The StudyWe analyzed 82 type and other culture collection strains encompassing all validly described staphylococcal species (n = 38) and subspecies (n = 21; according to the current List of Bacterial Names with Standing in Nomenclature, updated May 14, 2005) (7). Two strains of the recently proposed candidate species S. pettenkoferi (8) were added to complete the rpoB sequence reference database. Using this database, we analyzed 55 clinical staphylococcal isolates collected from human (n = 52) and animal (S. intermedius, n = 2; S. felis, n = 1) specimens; 6 of the human isolates exhibited the small-colony variant (SCV) phenotype. This strain collection was previously analyzed by the API ID 32 Staph and VITEK 2 systems (both obtained from bioMérieux, Marcy l'Etoile, France), partial 16S rDNA sequencing, chemotaxonomy, and riboprinting to determine species designation (3). The thermal cycling condition to amplify the partial rpoB gene (899 bp) was 35 cycles of denaturation at 94°C for 45 s (300 s for the first cycle), annealing (60 s at 52°C), and extension (90 s at 72°C, 600 s for the last cycle). The Staphylococcus-specific primers used for amplification and sequencing of rpoB are shown in Table 1. Sequencing reactions were performed in a total volume of 10 μL containing 0.5 μL premix (ABI Prism BigDye Terminator v3.0 Ready Reaction Cycle Sequencing Kit, Applied Biosystems, Darmstadt, Germany), 1.8 μL 400 mmol/L Tris-HCl, 10 mmol/L MgCl2, 10 pmol sequencing primer, and 2 μL polymerase chain reaction product. The sequencing products were purified by using the Centri-Sep Spin Columns (Princeton Separations, Adelphia, NJ, USA) and analyzed with the ABI Prism 3100 Avant Genetic Analyzer (Applied Biosystems) according to the manufacturer's instructions. For further analysis, nucleotides 1444–1928 (corresponding to S. aureus rpoB gene positions of the GenBank accession no. X64172) of the rpoB gene were used. The sequences were analyzed by using Ridom TraceEditPro version 1.0 software (Ridom GmbH, Würzburg, Germany). Staphylococcal partial rpoB reference sequences determined in this study were deposited in GenBank under accession nos. DQ120729–DQ120752.
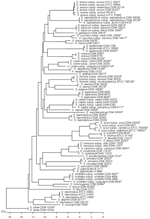
Partial rpoB sequences were determined for 82 culture collection strains and 55 clinical isolates. All staphylococcal type strains were distinguishable by rpoB; the only exception was the S. equorum subspecies that shared the same sequence (Appendix Figure). The mean pairwise distance of all type and other culture collection strains exhibiting a unique rpoB sequence (n = 68) was 13.7% (range 0%–21.4%) and the standard deviation was 3.3%. When assuming a normal distribution for the distances and choosing a reporting criterion >94.0%, the similarity for a distinct species correlates with a statistical error probability of 1.0% (9). The definitive identification of 55 clinical isolates and the rpoB gene sequence similarity search results are shown in Table 2. At the species level, the correct species designation for all 55 clinical isolates was made by rpoB sequence similarity search (sequence similarity >94.0%). Of 21 clinical isolates belonging to species currently divided into subspecies, 17 isolates were correctly identified to the subspecies level. Subspecies identification for isolates M26 and M53 was unsuccessful by rpoB or partial 16S rDNA sequencing, riboprinting, and chemotaxonomy (data not shown). Only isolates M20 and M39 were misidentified by rpoB sequencing as S. saprophyticus subsp. saprophyticus instead of subsp. bovis. ConclusionsOur previous study demonstrated the superiority of sequence-based methods over phenotypic approaches using the API ID 32 Staph and VITEK 2 systems (3). The advantage of a sequence-based method became most evident when differentiating isolates with the SCV phenotype, in which the API ID 32 Staph and VITEK 2 systems misidentified 2 and 4 isolates, respectively. When both sequence-based approaches used were compared, rpoB sequencing was superior to partial 16S rDNA identification. Although the 16S rDNA procedure differentiated 50 (90.9%) of all tested clinical isolates at species level, rpoB identified 100%. Therefore, if an unknown organism needs to be identified, 16S rDNA sequencing is the method of choice because of the availability of universal primers (10). However, if the genus is already known, the rpoB method should be used. Compared with other published molecular probes, rpoB showed the highest discriminatory power, e.g., hsp60 and sodA sequencing did not differentiate subspecies of S. carnosus, S. cohnii, S. hominis, S. schleiferi, or S. succinus (4,5). In a previous study, rpoB sequence-based identification of Staphylococcus species has been reported (6). However, a limited number of taxa were included, and the primers used were not appropriate to detect all staphylococcal subspecies. Sequencing of rpoB was also used to identify other bacterial species (11,12). A higher discrimination with rpoB sequencing compared with 16S rDNA sequencing has been demonstrated for the genera Corynebacterium (13) and Bacillus (14). DNA sequencing is a rapid alternative to biochemical and other phenotypic procedures for the differentiation of bacterial pathogens because of its decreased costs and increased automation (15). Thus, rpoB is a useful molecular target for differentiating staphylococcal isolates to the species and subspecies level.
References
Suggested citation
for this article: |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
||||||||||||
|
||||||||||||
|
|
|
EID Home | Top of Page | Ahead-of-Print | Past Issues | Suggested Citation | EID Search | Contact Us | Accessibility | Privacy Policy Notice | CDC Home | CDC Search | Health Topics A-Z |
||
|
This page
posted January 24, 2006 |
||
|
Emerging
Infectious Diseases Journal |
||