|
The mysterious origin of strange growths on the stems, leaves, flowers and roots of plants have intrigued naturalists for centuries. Called galls or hypertrophies, these tumorous (neoplasmic) outgrowths develop from rapid mitosis and morphogenesis of plant tissues and come in an astounding array of colors, shapes and sizes. Galls may be smooth, spiny or fuzzy, and resemble everything from marbles and ping-pong balls to dunce caps, saucers and sea urchins. Many galls provide the food and brooding structure for various species of harmless insects. Because of the general nature of galls, many authors refer to any swollen structure bearing insects as galls. This is not only incorrect, but it detracts from the marvelous evolution of some remarkable plant species which have literally developed little "condos" to house their special symbiotic insects. These help-mate insects provide a vital service to their host plant in the form of pollination or protection in a highly competitive environment where these plants could otherwise not survive.

|
Swollen stem galls on the branchlets of a canyon live oak (Quercus chrysolepis) caused by a cynipid gall wasp (Andricus spectabilis?).
|
2. Destructive Galls Caused By Fungi, Bacteria & Gall Midges
For countless centuries plants have been forming galls to ward off the attacks of bacteria, fungi, mistletoe, mites, nematodes, viruses, and insects. Remarkable fossil galls have been found on extinct seed ferns and conifers dating back more than 200 million years. Although most galls probably do not harm the host too much, they do involve some form of parasitism and generally do not benefit the host. Gall midges include a number of minute flies, some of which attack valuable crops such as clover, alfalfa and grains. One of the most destructive is the Hessian fly (Phytophaga destructor), possibly brought to America by mercenary Hessian troops serving with the British during the Revolutionary War. Masses of the larvae destroy the wheat crop by feeding between the leaf sheath and the stem of wheat.

|
Unsightly peach leaf curl caused by the parasitic fungus Taphrina deformans, a member of the fungal Division Eumycota, class Ascomycetes. Because this fungal infection causes tumorous swellings on leaves, it is often referred to as a gall. If not controlled, it can cause serious damage to orchard trees.
|
Like tumorous growths, some galls can be very destructive to the host plant. In fact, crown gall (caused by the bacterium Agrobacterium tumefaciens) and peach leaf curl (caused by the fungus Taphrina deformans, Class Ascomycetes) can cause serious damage to orchard trees. Crown galls are especially interesting because the plasmids of this bacterium (small, circular DNA molecules) contain genetic information that may become incorporated into the nuclear DNA of the infected host cells. These plasmids are referred to as tumor-inducing or Ti plasmids. The Ti plasmid DNA contains genes which cause the host cells to divide uncontrollably. In a sense, the plasmids are like floppy disks containing files or "genes" which are loaded into the host DNA. This explains why these "plant tumors" continue to grow even when the bacteria are eliminated. Ti plasmids are a powerful tool for genetic engineering in plants because the tumor-inducing genes can be replaced with beneficial genes. Genetically-engineered plasmids can then be used to transfer the beneficial genes into the host DNA. For example, bacterial genes for insecticidal proteins have been transferred into tomato plants, resulting in transgenic plants that are resistant to ravenous lepidopteran larvae. The protein toxin actually kills the caterpillars feeding on the plant.

|
Clear evidence of crown gall at the base of a deceased apple tree. Note the enlarged swellings on the roots. This plant disease is caused by the soil bacterium (Agrobacterium tumefaciens) which infects healthy plants, especially if they are wounded.
|

|
Massive crown gall at the base of an adventitious stem of an apple tree. Note the enlarged swellings on the roots. This plant disease is caused by the soil bacterium (Agrobacterium tumefaciens).
|

|
Three unsightly eruptions of crown gall at the base of a sick rose plant. This serious disease of roses is caused by the soil bacterium Agrobacterium tumefaciens.
|
3. Galls Caused By Mistletoe
The exact cause of gall formation varies with different plant species. In the case of certain parasitic fungi and mistletoes, the host plant tends to produce a mass of woody tissue surrounding the infection. This is especially evident on the stems and branches of oaks where the specialized mistletoe root system (called haustorium) has penetrated the host tissue. Long after the soft-tissued mistletoe has died and withered away, the persistent gall remains on the limbs. Old patriarch oaks may have huge galls several feet in diameter, battle scars from mistletoe attacks many decades before. Some tropical mistletoes leave perfect attachment imprints on the host stem long after they have rotted away and fallen off. These club-shaped malformations of the host wood, intricately sculptured by the impressions of the mistletoe haustorium, are called "wood roses." They are a perfect cast of the parasite's haustorial attachment. In Mexico and Bali, wood roses are carved into all sorts of beautiful figures, including fish, birds and reptiles. Other unusual mistletoe galls called "witches' brooms" are occasionally found on cone-bearing trees of the Pacific northwest. These striking, unmistakable galls are often caused by the dwarf mistletoe (Arceuthobium) and result in dense clusters of shoots and stems near the top of the tree.
4. Fasciated Stem Galls
Another gall-like structure is called a fasciation. Sometimes the flower stalks and terminal branches of wildflowers and shrubs become enlarged and flattened into a bizarre, fan-shaped structure quite unlike the normal stem. Fasciations occur on a wide variety of plants, including sagebrush (Artemisia), chocolate lilies (Fritillaria), evening primroses (Oenothera) and a tall roadside weed called mullein (Verbascum thapsus). Although some fasciated stems may be a genetic malformation (as in certain cristate cacti), they may also be gall-like in nature and are caused by insects, mites and nematodes.
5. Bizarre Galls Caused By Fungi
Some of the most unusual and remarkable galls are caused by fungal mycelia that permeate the host tissue. The familiar reddish balls called "cedar apples" which decorate many species of junipers are actually galls caused by rust fungi of the genus Gymnosporangium. In the spring, the galls are covered with radiating gelatinous spikes bearing thousands of special spores called teleutospores. These spores must infect nearby apple (Malus), crab apple and hawthorn (Crataegus) trees in order to complete their life cycle. It is generally wise not to plant junipers close to your apple orchard if you prefer apples to cedar apples.

|
Fungus stem gall or "poop gall" on choke cherry (Prunus virginiana) in Montana. The swollen stems are caused by the fungus Dibotryon morbosum.
|
The most extraordinary fungus galls are caused by rust fungi of the genus Puccinia. In fact, these abnormal growths are so bizarre that some authorities might be hesitant to call them galls. Certain Puccinia species are known to attack perennial rock cresses (Arabis) of the Rocky Mountains, causing the mustards to produce flower-like, erect, leafy stalks called "pseudoflowers." The bright rosettes of showy leaves are covered with droplets of sweet, fragrant nectar and fungal reproductive sex cells called spermatia. Attracted by the sweet honeydew secretion, insects transfer the "male" and "female" spermatia to other pseudoflowers--thus enabling sexual reproduction for the fungus. The effects of this strange twist in pollination strategy on flowering plants is currently being studied by pollination biologists.
6. Galls Caused By Insects

|
A canyon live oak (Quercus chrysolepis) stem gall in the palm of a hand. A: The larva inside of the gall; B: A minute, adult, cynipid gall wasp (Andricus spectabilis?).
|

|
A "tootsie-pop" gall on the stem of creosote bush (Larrea tridentata) caused by the gall midge (Asphondylia auripila).
|
Probably most of the galls that you encounter are actually caused by minute insects, particularly flies, wasps and aphids (homopterans). Sometimes the gall-forming insects are prolific. On a recent survey of a 50 acre disturbed grassland area in southern California, practically every individual of sand aster (Corethrogyne filaginifolia) had a swollen stem tip gall covered with a dense rosette of leaves. A few examples of the diversity of galls in California include the creosote bush "tootsie-pop" gall caused by the gall midge (Asphondylia auripila), the spruce pineapple gall caused by the spruce gall aphid (Adelges cooleyi), the ubiquitous oak apple gall caused by the California gall wasp (Andricus californicus), the oak saucer-gall caused by the gall wasp (Andricus gigas), the spined-turban gall caused by the gall wasp (Antron douglasii), and the unusual Oregon oak "ping-pong" ball gall caused by the gall wasp (Cynips maculipennis). The largest percentage of these insect galls are found on plants in the rose, juniper, willow, sunflower and oak families. According to R.A. Russo (Plant Galls of the California Region, 1979), almost half of the known galls in America occur on oak trees (Quercus). In fact, many of the wasps causing these galls belong to a single family of gall wasps, the Cynipidae. Another native tree that for some reason is typically heavily infested with insect galls is the western hackberry (Celtis reticulata). On a ridge high above the headwaters of the San Diego River (interior San Diego County), old, gall-covered western hackberry trees form grotesque images against the late afternoon sky.

|
An "oak apple" gall caused by the California gall wasp (Andricus californicus).
|
Most of these unusual and colorful insect galls are relatively harmless to their host plants, and the larvae inside are often eaten by a variety of birds and rodents. Some galls contain a very high content of bitter tannins, and are probably unpalatable to predators. In fact, oak galls were gathered in Europe and Asia Minor as a source of tannin used for converting animal skin into leather. Gallic acid, first isolated from oak leaf galls by the Swedish chemist Karl Scheele in 1786, is also used in tanning, dyes, inks, photographic developers and antioxidants.

|
A saucer gall on the leaf of a scrub oak (Quercus cornelius-mulleri) caused by the gall wasp Andricus gigas.
|
7. The Initiation Of Insect Galls
The most fascinating galls are sometimes referred to as "controlled galls" because they grow into a specific size, shape and color that is characteristic of a particular species of wasp. Gall formation begins when a female gall wasp injects her eggs into a bud, leaf or stem. Upon hatching from the eggs, the hungry larvae begin feeding on the host tissue surrounding them. The plant's defensive reaction to this intrusive mechanical or chemical irritation is to isolate the toxins or activities of the invader in a tough, tumorous mass of tissue called a gall. Ironically, in doing so the plant provides food and shelter for the developing ravenous larvae. After completing their growth and metamorphosis, often many months later, the adult wasps escape by chewing a circular exit tunnel through the wall of the gall. The precise mechanism by which different species of wasps produce such remarkably unique galls is still being debated by cecidologists (people who study galls).
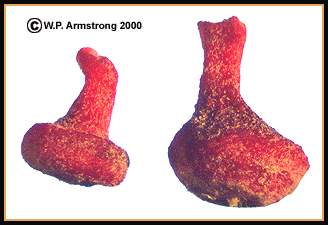
|
Red cone galls from the leaf of a valley oak (Quercus lobata) caused by the gall wasp Andricus kingii.
|
The initiation of insect galls is typically associated with oviposition by the adult (e.g. sawflies, gall wasps and beetles) or by the feeding of early larval stages (e.g. midges, moths and aphids). Depending on the type of insect and whether it has chewing or piercing-sucking mouthparts, a variety of salivary fluids may be injected into the plant tissue. According to K. Hori (Biology of Insect-Induced Galls, Chapter 10, 1992), salivary fluids of hemipterans (bugs) and homopterans (aphids) may include amino acids, auxins (IAA) and various plant digesting enzymes such as pectinases, cellulases and proteases. The precise mechanism by which these chemicals induce cell division and morphogenesis is very complicated and varies with different species and even with different types of plant tissue. An increased concentration of certain plant hormones in the gall tissue may also be important for the morphogenesis of some galls. A fascinating summary of research on insect galls is presented in Biology of Insect-Induced Galls, edited by J.D. Shorthouse and O. Rohfritsch, Oxford University Press, 1992.

|
Striking "ping-pong" ball galls of the minute gall wasp Cynips maculipennis on the leaves of Oregon oak (Quercus garryana).
|

|
Close-up view of the undersides of blue oak leaves (Quercus douglasii) showing several fuzzy galls caused by the cynipid gall wasp (Andricus brunneus).
|

|
Southern live oak (Quecus virginiana), an evergreen oak native to the eastern United States. The fuzzy caterpillarlike galls on the underside of the leaf are caused by the gall wasp Andricus, possibly A. chrystallinus. This gall is common on California white oaks, including the blue oak (Q. douglasii), leather oak (Q. durata) and Oregon oak (Q. garryana). This wasp apparently also likes the Virginia oaks in the Palomar College Arboretum.
|

|
Distinctive chinquapin galls on the bush chinquapin (Chrysolepis sempervirens) in the Sierra Nevada of California. The galls are caused by the cynipid wasp (Dryocosmus castanopsidis).
|
8. The Spruce Pineapple Gall

|
Spruce pineapple galls on the swollen, needle-bearing branchlets of Engelmann spruce (Picea engelmannii) and Colorado blue spruce (P. pungens) in Montana. The galls are caused by an aphid-like insect (Adelges cooleyi) of the order Homoptera that invades the needle-bearing branch tips. In California, this insect also attacks the sitka spruce (P. sitchensis), weeping spruce (P. breweriana) and several cultivated trees, including the oriental spruce (P. orientalis) and Norway spruce (P. excelsa). The tiny insects escape from the galls through lip-like slits above each needle. This insect has a complex life cycle that involves two genera of host trees, including several species spruce (Picea) and Douglas fir (Pseudotsuga meziesii).
|

|
Tiny adult females emerge from the pineapple galls in late summer. Each female has an aphid-like body with sucking mouthparts and two pairs of transparent wings.
|
The winged females fly to nearby Douglas fir trees where they lay eggs on the needles. Several generations of cotton-like woolly aphids are produced. Late in summer, some of the woolly aphids develop wings and fly back to the spruce trees to deposit eggs, which produce the overwintering population. Other wingless aphids remain on Douglas fir trees where they also produce overwintering forms. No galls are formed on the Douglas fir trees. According to D.J. Borror and D. M. Delong (An Introduction to the Study of Insects, 1964), only female generations are produced, a phenomenon known as parthenogenesis. Swarming masses of these minute insects may be observed in northern Montana during late summer and fall.
9. California's Amazing Jumping Galls
Certainly one of the most interesting galls are the "jumping galls" of California. Jumping galls are attached to the leaves of several native oaks in California's Sacramento Valley, including the valley oak (Quercus lobata), blue oak (Q. douglasii ) and Oregon oak (Q. garryana). During favorable years, the undersides (and uppersides) of each leaf contains dozens of galls. When multiplied by the hundreds of thousands of leaves per tree, this accounts for the millions of galls that fall to the ground beneath these oaks, like a shower of tiny BB-shaped bird seed. In fact, some valley residents become dismayed when the galls cover their patios, sidewalks and driveways. Each gall is inhabited by a tiny cynipid gall wasp appropriately named the "jumping oak gall wasp" (Neuroterus saltatorius), formerly named Cynips saltatorius. Since the gall consists of a single cavity or chamber occupied by a single wasp it is termed monothalamous. And since the galls break away from the leaves, they are called "detachable galls." When the minute galls fall to the ground they begin hopping about like fleas. Like jumping beans, the larva inside is active during the summer months, but ceases its activity by late summer and fall when it changes into a pupa. And like jumping beans under jumping bean shrubs, the sound of thousands of jumping galls in the leaf litter beneath oaks resembles the patter of rain drops falling on dry leaves.
10. Mexican Jumping Beans: Not Galls Or Beans
There are many examples of swollen, gall-like plant structures containing insects that are not galls at all. In some cases the larvae of these insects are simply seed predators that consume the contents of an ovary or seed capsule. After pollination and fertilization, the nutritive seed tissue develops with or without the presence of their symbiotic insects. The inflated, papery Mexican jumping beans that roll around by seemingly perpetual motion are actually sections (carpels) of seed capsules of the jumping bean shrub (Sebastiana pavoniana) which contain the larva of the jumping bean moth (Laspeyresia saltitans). A similar larva also inhabits the seed capsules of the Arizona jumping bean shrub (Sapium biloculare). Upon hatching, the larva bores into the green, immature capsule while attached to the host shrub and proceeds to feed on the seed tissue inside. Luckily for the shrub, some carpels do not contain the ravenous larva and develop a mature seed.
11. The Fig Gall Controversy
Although some authors refer to the "fruit-like" structures on wild, fig trees (Ficus) as galls, they are actually specialized structures called syconia bearing minute male and female flowers on the inside. A tiny female wasp enters an opening (ostiole) on the syconium to pollinate the flowers and lay her eggs inside the short-style female flowers. She inserts her ovipositor down the stylar canal and deposits an egg inside the ovary of each short-style flower. According to I.J. Condit (The Fig, 1947), oviposition injures the stylar canal, thus inhibiting pollen tube growth and fertilization in short-style flowers. Because her ovipositor is too short, the fig wasp is unable to oviposit inside the long-style flowers. The latter flowers each develop a seed (with an embryo and endosperm) by normal pollination and double fertilization. Although there is considerable disagreement in the literature, many authors continue to describe the short-style flowers as "gall flowers," presumably because they are commonly occupied by a developing male or female fig wasp; however, they are fully capable of producing normal seed-bearing drupelets, and in this respect are no different from long-style flowers. In 2-3 months a new crop of male and female wasps emerge from the short-style flowers. After insemination by males, the fertile female wasps pack their pollen baskets (corbiculae) with pollen and exit the syconium. These syconia serve as "wasp condos" and seed-bearing structures for populations of wasps that pollinate the fig flowers in one of nature's most remarkable and complex symbiotic cycles.
The "gall controversy" of fig flowers is complicated because food tissue (endosperm) for the developing larva may be initiated parthenogenetically (without pollination and fertilization), possibly by a mechanical or chemical stimulus during oviposition. In this case the flower functions like a minute gall, except their is no apparent tissue malformation as in typical insect galls. Functional male caprifigs of Ficus carica produce three crops of syconia per year: the summer profichi, fall mammoni and overwintering mamme that mature the following spring. Only the profichi crop produces pollen, and this is used to pollinate the Calimyrna orchards of California's Central Valley. The receptive mamme and profichi syconia are not pollinated, so endosperm tissue to nourish the wasp larvae in these crops must be initiated parthenogenetically. Whether they contain a mature seed or a wasp, the short-style flowers of caprifigs appear virtually identical in structure, and do not fit the definition of a typical gall. However, proponents of gall flower terminology argue that when the wasp induces the formation of nutritive endosperm tissue, the ovary interior is literally transformed into a minute gall.
Another case for wasp-induced "gall-like" endosperm tissue is the sycomore (sycamore) fig (Ficus sycomorus) of the Middle East. According to J. Galil (Endeavor, Vol 1, 1977), oviposition by the nonpollinator wasp Sycophaga sycomori results in the development of parthenogenetic endosperm tissue which the larva feeds upon. The galled endosperm tissue develops through proliferation of nucellar cells surrounding the embryo sac. Since there is no pollination, the syconia are seedless. However, in its native habitat of eastern Central Africa and Yemen, F. sycomorus is pollinated by the symbiotic wasp Ceratosolen arabicus, and the long-style flowers produce seeds. The story is even more complicated because without pollination the sycomore fig typically drops its immature syconia. Oviposition and the presence of nonpollinator wasp larvae not only initiate the development of endosperm tissue, but also the enlargement and ripening of the syconium containing wasp-bearing drupelets without pollination--a condition known as parthenocarpy. Since the normal course of events is to abort unpollinated syconia, the entire syconium could be viewed as a gall occupied by nonpollinator wasps. The sycomore fig was carried north to Egypt by 3000 B.C., and on to Israel, Lebanon and Cyprus. Early farmers in these regions learned how to induce parthenocarpy in sycomore figs by gashing them with a knife. Within 3-4 days the hard, green syconia enlarge and become sweet and fleshy, long before any possible wasp inhabitants are sufficiently developed to damage the crop. In fact, some biblical scholars think the phrase "gatherer of sycomore fruit" (Amos 7:14) actually means "piercer of sycomore fruit." According to J. Galil (Economic Botany Vol. 22, 1968), gashed figs produce ethylene gas which hastens the ripening process. It is interesting to note that in Israel today there is a fully parthenocarpic variety of sycomore fig that does not require gashing or wasps. The syconia ripen on the tree without any external stimulus, as in the popular black mission, kadota and brown turkey varieties of F. carica.
12. The Desert Trumpet Gall Controversy
Most floras of California and the southwest region (including the recent Jepson Manual, 1993) list two varieties of the striking wildflower called desert trumpet (Eriogonum inflatum): Var. inflatum with conspicuous swollen stems and var. deflatum with normal slender stems. According to E.C. Jaeger (Desert Plants, 1941), wasps of the genus Onyerus sometimes use the hollow stems as a larder and brooding chamber. The female wasp drills a hole in the hollow stem, packs the cavity with dead or paralyzed insect larvae, and then lays her eggs on the abundant food supply. Jaeger's amazing observation would hardly constitute a gall; however, this subject is considerably more controversial. A.M. Stone and C.T. Mason, Jr. (Desert Plants Vol. 1, 1979) studied wild and greenhouse populations of both varieties and found that stem inflation was caused by larval feeding of gall insects, including a lepidopteran (Pyralidae) and a beetle (Mordellidae). They concluded that "the taxonomic recognition of the varieties inflatum and deflatum is not based on a genetic characteristic, but on a monstrosity, and has no validity."
The Desert Trumpet Riddle Solved At Last
According to the world authority on the genus Eriogonum, Dr. James L. Reveal of the University of Maryland (personal communication, 1998), the swollen stem of E. inflatum var. inflatum is due to high concentrations of carbon dioxide (CO2) in the solid stem and seems to be related to gas regulation. Although some insects utilize the swollen stem as a larder, the inflation is NOT caused by the larval feeding of gall insects. In fact, during very dry years many populations of var. inflatum will have no or poorly inflated stems. In northeastern Utah and adjacent Colorado, another species E. fusiforme occurs in the tens of millions in a good year, and all have inflated stems even when they are young (and there is no evidence that the inflated stems are caused by insects).
|
13. The Swollen-Thorn Acacia Gall Misnomer
Another undisputable gall misnomer concerns the inflated thorns on the African and Central American swollen-thorn acacias. The peculiar thorns are hollowed out and occupied by fiercely biting/stinging symbiotic ants (Pseudomyrmex ferruginea) that protect the trees from browsing herbivores and from epiphytic plants that might shade out the acacias. In the Central American Acacia collinsii, the ants actually clear away invasive seedlings around the base of the tree that might overgrow the acacia and block out vital sunlight. According to Daniel Janzen (Smithsonian Contributions to Botany Number 13, 1974), the swollen-thorn acacias lack the chemical and physical defenses of most other acacias to deal with their predators and competition, and the symbiotic ants have taken over this vital role. The acacias reward their little ant colonies with thorn "condos" to live in, carbohydrate-rich nectar from glands on the leaf stalks, and nourishing, protein-lipid Beltian bodies on the leaflet tips. These remarkable acacias grown in cultivation far away from their native habitat still produce their striking swollen thorns without the presence of their symbiotic ants. I have seen the Central American A. cornigera in San Diego County, complete with its swollen thorns and Beltian bodies, and no symbiotic ants to protect it. The next time you visit the San Diego Zoo, notice the inflated thorns of the whistling-thorn acacia (A. drepanolobium) near the exit gate. [In South Africa the hollowed-out thorns whistle when the wind blows across their entrance holes.] These amazing thorns are not galls, they are genetically-programmed structures that have evolved over millions of years of coexistence with ants. In South Africa the bulbous thorns of this acacia are occupied by stinging ants of the genus Crematogaster which protect the trees from destructive herbivores.
To the casual observer, galls may appear to be unsightly growths that mar the beauty of wild and ornamental plants. However, the complex interactions between the gall-maker organism and its host plant are truly remarkable and may have valuable agricultural applications. For example, by replacing the plasmid DNA of infectious crown gall bacteria with beneficial genes, new "transgenic plants" can be produced which are resistant to insects, herbicides and viruses. The precise mechanism of gall formation may also have valuable medical implications, particularly in understanding the rapid growth of renegade tumor cells.
14. WAYNE'S WORD Links About Galls:
15. WAYNE'S WORD Links About Figs & Fig Wasps:
15. References About Galls
- Armstrong, W.P. 1995. "To Be Or Not To Be A Gall." Pacific Horticulture 56: 39-45.
- Armstrong, W.P. 1995. "A Galling Puzzle." Zoonooz 68 (3): 26-31.
- Armstrong, W.P. 1988. "The Calimyrna Fig and Its Wasp." California Garden 79: 135-138.
- Condit, I.J. 1947. The Fig. Chronica Botanica Co., Waltham, Mass.
- Cowen, R. 1990. "Parasite Power." Science News 138: 200-202.
- Galil, J. 1977. "Fig Biology." Endeavour 1: 52-56.
- Galil, J. 1968. "An Ancient Technique for Ripening Sycomore Fruit in East-Mediterranean Countries." Economic Botany 22: 178-190.
- Galil, J. 1967. "Sycomore Wasps From Ancient Egyptian Tombs." Israel Journal of Entomology II: 1-10.
- Galil, J. and D. Eisikovitch. 1974. "Further Studies On Pollination Ecology in Ficus sycomorus. II. Pocket Filling and Emptying by Ceratosolen arabicus." Magr. New Phytol.. 73: 515-528.
- Hickman, J.C. (Editor). 1993. The Jepson Manual of Higher Plants of California. University of California Press, Berkeley.
- Hogue, C. L. 1993. Insects of the Los Angeles Basin. Natural History Museum of Los Angeles County, Los Angeles, California.
- Jaeger, E.C. 1941. Desert Wildflowers. Stanford University Press, Stanford, California.
- Janzen, D.H. 1974. Swollen-Thorn Acacias of Central America. Smithsonian Contributions To Botany Number 13. Smithsonian Institution Press.
- Kuijt, J. 1969. The Biology of Parasitic Flowering Plants. University of California Press, Berkeley.
- Leach, F.A. 1923. "Jumping Seeds: Plant Growths That Hop About Like Fleas." Natural History 23: 295-300.
- Pavlik, B.M., P. C. Muick, S.G. Johnson and M. Popper. 1991. Oaks in California. Cachuma Press, P.O. Box 560, Los Olivos, California.
- Rosenthall, S.S. and C.S. Koehler. 1971. "Heterogeny in Some Gall-Forming Cynipidae (Hymenoptera) With Notes on the Biology of Neuroterus saltatorius." Annals of The Entomological Society of America 64: 565-570.
- Rosenthall, S.S. and C.S. Koehler. 1971. "Intertree Distributions of Some Cynipid (Hymenoptera) Galls on Quercus lobata." Annals of The Entomological Society of America 64: 571-574.
- Russo, R. A. 1979. Plant Galls of the California Region. Boxwood Press, Pacific Grove, California.
- Russo, R. A. 2006. Field Guide To Plant Galls of California and Other Western States. University of California Press, Berkeley, California.
- Shorthouse, J.D. and O. Rohfritsch (editors). 1992. Biology of Insect-Induced Galls. Oxford University Press.
- Stone, A.M. and C.T. Mason, Jr. 1979. "A Study of Stem Inflation in Wild Buckwheat, Eriogonum inflatum." Desert Plants 1 (2): 77-81.
|
|