|
| |
||
| |
|||||||||||||||||
|
|||||||||||||||||
|
|||||||||||||||||
|
EID Home | Ahead of Print | Past Issues | EID Search | Contact Us | Announcements | Suggested Citation | Submit Manuscript
|
|
Dispatch Human Escherichia coli O157:H7 Genetic Marker in Isolates of Bovine OriginJeffrey T. LeJeune,*
Escherichia coli O157 is recognized worldwide as an important cause of diarrheal disease, which in some patients is followed by hemolytic uremic syndrome and death (1). A primary virulence factor of this pathogen is the prophage-encoded Shiga toxin (2). Greater Shiga toxin production per bacterium is associated with increasing severity of human disease (3,4). Because of its location in the phage genome, the stx-gene variant dubbed stx2 is under similar regulatory control as other phage late-genes, as it is governed by the interaction of the transcription antiterminator Q with the late promoter PR´ (5). Although cattle and other ruminants appear to be the natural reservoir for E. coli O157 and other Shiga toxin–producing E. coli (STEC), only a small fraction of STEC serotypes routinely present in cattle are frequently isolated from human patients. Mounting evidence suggests that considerable genetic, phenotypic, and pathogenic diversity exists among these pathogens (6–8). Furthermore, genetic subtypes or lineages of E. coli O157 do not appear to be equally distributed among isolates of bovine and human origin (7). The purpose of this study was to examine the distribution of specific sequences upstream of the stx2 gene among E. coli O157:H7 of human and bovine origin, along with corresponding magnitudes of Shiga toxin production. The StudyA total of 158 stx2-encoding E. coli O157:H7 isolates were assayed, 91 isolates of bovine origin and 67 originally isolated from ill persons (Appendixes 1 and 2). All isolates demonstrated unique banding patterns on pulsed-field gel electrophoresis (PFGE). For polymerase chain reaction (PCR) analysis, 5 µL of DNA obtained from boiled stationary-phase bacteria was added to a 50-µL PCR master mix containing a final concentration of 1.5 (Q933) or 2.5 (Q21) mmol MgCl2, 200 µmol/L each deoxynucleoside triphosphate, 1 U Taq polymerase, 0.6 pg/µL of primer 595 (5´-CCGAAGAAAAACCCAGTAACAG-3´) (9), and 0.6 pg/µL of either primer Q933 (5´-CGGAGGGGATTGTTGAAGGC-3´;QStxf) (9) or primer Q21 (5´-GAAATCCTCAATGCCTCGTTG-3´; this study). PCR consisted of an initial denaturation at 94°C for 5 min; 30 cycles of 94°C for 30 s, 52°C (Q933) or 55°C (Q21) for 1 min, and 72°C for 1 min; and a final 10-min extension step at 72°C. E. coli strain 933 or FAHRP88 was used as a positive control and master mix alone as a negative control. All PCR products were separated by gel electrophoresis (100 V) in 1% agarose gels, stained with ethidium bromide, and visualized by using UV illumination. Shiga toxin production was determined by using a commercially available enzyme-linked immunosorbent assay (ELISA) kit (Premiere EHEC, Meridian Diagnostics, Cincinnati, OH). Briefly, log-phase cells from Luria-Bertani broth enrichments were diluted to 0.6 optical density (OD) at 600 nm, subsequently pelleted, resuspended in phosphate-buffered saline, and induced by exposure to UV light (240 nm) for 3 s (10). A 1:9 volume of a 10x concentrate of brain heart infusion broth was added to each culture and shaken at 37°C for 2.5 h. Replicate cultures that were not exposed to UV light (noninduced controls) were maintained at 4°C. Two hundred microliters of each induced and noninduced enrichment was subsequently used as the specimen in the EHEC ELISA, as described (11). OD results were recorded for each isolate both with and without UV induction. The relative change in Shiga toxin production after induction was calculated for each isolate; (ODinduced)/ODnoninduced). E. coli O157 (EDL933) and a toxin-negative control isolate were assayed as positive and negative controls each time the assay was repeated.
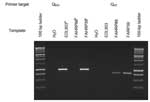
E. coli O157 isolates were classified on the basis of the presence or absence of bands of the predicted size on the Q933-595 and Q21-595 PCR reactions (Figure). A chi-square test was used to determine whether different PCR genotypes were equally distributed among isolates of bovine and human origin. Likewise, a chi-square test was used to assess the equality of distribution of PCR genotypes among bovine isolates from different countries. One-way analysis of variance for nonparametric data (Kruskal-Wallis test) was used to identify differences in ranked-transformed toxin production among noninduced and induced E. coli O157 isolates as well as to determine significant differences in the percent increase in toxin following induction. Previously, Kim et al. described a nonrandom distribution of E. coli O157 subtypes among cattle and humans by using an octamer-based genome-scanning method (7). We tested several of the isolates that had been previously characterized. Nine had been previously identified as belonging to the lineage I genotype and seven isolates as belonging to the lineage II genotype. We found that all nine lineage I isolates consistently amplified the Q933 target, regardless of species of origin. All four bovine isolates classified as lineage II by Kim et al. amplified the Q21 target. One lineage II human isolate (NE015) amplified the Q933 target, and another lineage II isolate (NE037) produced no amplicons in either PCR reaction. One human isolate classified as lineage II (ATCC 43889) amplified both target sequences, presumably because of polylysogeny. The distribution of the specific Q-gene alleles found upstream of the prophage stx region among bovine isolates may have a geographic component. The distribution of E. coli O157 phage genotypes collected from healthy cattle from diverse geographic areas is consistent with the variable incidences of human disease in different countries (Table 1). For example, six (75%) of eight Scottish bovine isolates examined amplified the Q933 target, the same target that is frequently present in human isolates of human disease origin. Scotland reports some of the highest incidence rates of human E. coli O157–related diseases and hemolytic uremic syndrome (13). In contrast, none of the seven Australian E. coli O157 bovine isolates amplified the 1750-bp fragment. Contrary to the situation in Scotland and the United States, E. coli O157 infection of humans is rarely reported in Australia (14). ConclusionsThe Q933 gene target was more commonly identified in human disease–associated strains of E. coli O157 than from strains of bovine origin. Amplification of the Q933 target, either alone or in combination with amplification of the Q21 target from the same isolate, was identified in 60 (9%) of 66 (55/66 alone and 5/66 in combination with Q21; 1 isolate amplified neither target) compared to 40 (44%) of 91 (32/91 alone, and 8/91 in combination with Q21) of bovine isolates (p < 0.001). Furthermore, these genetic subtypes were nonrandomly distributed among the E. coli O157 isolates of bovine origin obtained from different countries (p < 0.05) (Table 1). These limited data suggest that the distribution of E. coli O157 strains in cattle may differ between countries or regions, thereby providing an explanation for geographic differences in the incidence of human E. coli O157 infection. More isolates from cattle need to be analyzed with these methods to better characterize the E. coli O157 in the bovine reservoir of each country. A positive reaction with the Q933 target was significantly associated with higher OD results on the Shiga toxin ELISA (both noninduced and induced) and higher-fold increases in toxin production following induction than isolates amplifying the Q21 target alone (p < 0.0001) (Table 2). Despite these differences, we did not identify any clinical associations between the magnitude of Shiga toxin production and severity of human disease could be identified in this study. Other, non–Shiga toxin–related virulence factors and host susceptibility are also believed to play essential roles in the outcome of clinical STEC infections. The Q933-negative isolates obtained from human disease might have lost this Q933-containing prophage by the time of isolation, or these isolates might have been recovered from patients also infected with STEC containing Q933-type prophage (15). Whether specific Q-gene alleles directly correlate with the magnitude of Shiga-toxin production or whether other (unstudied) factors within the phage lytic cascade genetically linked to specific Q alleles instead are responsible for the magnitude of toxin production is not known. The antiterminator Q, the protein product of the Q gene, and PR´, the late promoter, are reputed to be involved in regulating phage late-genes and, because of the location of PR´ in prophage genome, of Shiga toxin production as well (5). In E. coli O157 phage 933W (GenBank no. 9632466) and E. coli O157 stx2vhd (GenBank no. 15718404), the 359-bp sequence immediately upstream of the stx2 gene is nearly identical (>95% nucleotide identity). However, further upstream of this area of identity, DNA sequences differ significantly. In E. coli O157 933W, this gene is identified as the antiterminator Q gene. In contrast, in E. coli O157 stx2vhd this area is occupied by a gene with >95% sequence identity with the antiterminator Q gene of bacteriophage 21 (gi 4539472). The Q gene of bacteriophage 21 does not share DNA sequence homology with the Q gene of bacteriophage 933W, and only 36% predicted amino acid homology. Since the Q gene is reputed to play an important role in regulating toxin production, our results provide a plausible explanation (differential regulation of Shiga toxin production) of why certain E. coli O157 genotypes are more commonly isolated from human patients (7). Acknowledgments
References
|
||||||||||||||||||||||||||||||
|
|
||||||||||||
|
||||||||||||
|
|
|
EID Home | Top of Page | Ahead-of-Print | Past Issues | Suggested Citation | EID Search | Contact Us | Accessibility | Privacy Policy Notice | CDC Home | CDC Search | Health Topics A-Z |
||
|
This page posted July
2, 2004 |
||
|
Emerging
Infectious Diseases Journal |
||