
Dairy Chemistry and Physics

Composition and Structure
Physical Properties
The role of milk in nature is to nourish and provide immunological protection for the mammalian young. Milk has been a food source for humans since prehistoric times; from human, goat, buffalo, sheep, yak, to the focus of this section - domesticated cow milk (genus Bos). Milk and honey are the only articles of diet whose sole function in nature is food. It is not surprising, therefore, that the nutritional value of milk is high. Milk is also a very complex food with over 100,000 different molecular species found. There are many factors that can affect milk composition such as breed variations (see introduction, cow to cow variations, herd to herd variations - including management and feed considerations, seasonal variations, and geographic variations. With all this in mind, only an approximate composition of milk can be given:
- 87.3% water (range of 85.5% - 88.7%)
- 3.9 % milkfat (range of 2.4% - 5.5%)
- 8.8% solids-not-fat (range of 7.9 - 10.0%):
- protein 3.25% (3/4 casein)
- lactose 4.6%
- minerals 0.65% - Ca, P, citrate, Mg, K, Na, Zn, Cl, Fe, Cu, sulfate, bicarbonate, many others
- acids 0.18% - citrate, formate, acetate, lactate, oxalate
- enzymes - peroxidase, catalase, phosphatase, lipase
- gases - oxygen, nitrogen
- vitamins - A, C, D, thiamine, riboflavin, others
The following terms are used to describe milk fractions:
- Plasma = milk - fat (skim milk)
- Serum = plasma - casein micelles (whey)
- solids-not-fat (SNF) = proteins, lactose, minerals, acids, enzymes, vitamins
- Total Milk Solids = fat + SNF
Not only is the composition important in determining the properties of milk, but the physical structure must also be examined. Due to its role in nature, milk is in a liquid form. This may seem curious if one takes into consideration the fact that milk has less water than most fruits and vegetables. Milk can be described as:
- an oil-in-water emulsion with the fat globules dispersed in the continuous serum phase
- a colloid suspension of casein micelles, globular proteins and lipoprotein partilcles
- a solution of lactose, soluble proteins, minerals, vitamins other components.
Looking at milk under a microscope, at low magnification (5X) a uniform but turbid liquid is observed. At 500X magnification, spherical droplets of fat, known as fat globules, can be seen. At even higher magnification (50,000X), the casein micelles can be observed. The main structural components of milk, fat globules and casein micelles, will be examined in more detail later.

The fat content of milk is of economic importance because milk is sold on the basis of fat. Milk fatty acids originate either from microbial activity in the rumen, and transported to the secretory cells via the blood and lymph, or from synthesis in the secretory cells. The main milk lipids are a class called triglycerides which are comprised of a glycerol backbone binding up to three different fatty acids. The fatty acids are composed of a hydrocarbon chain and a carboxyl group. The major fatty acids found in milk are:
Long chain
- C14 - myristic 11%
- C16 - palmitic 26%
- C18 - stearic 10%
- C18:1 - oleic 20%
Short chain (11%) - C4 - butyric*
- C6 - caproic
- C8 - caprylic
- C10 - capric
* butyric fatty acid is specific for milk fat of ruminant animals and is resposible for the rancid flavour when it is cleaved from glycerol by lipase action.
Saturated fatty acids (no double bonds), such as myristic, palmitic, and stearic make up two thirds of milk fatty acids. Oleic acid is the most abundant unsaturated fatty acid in milk with one double bond. While the cis form of geometric isomer is the most common found in nature, approximately 5% of all unsaturated bonds are in the trans position as a result of rumen hydrogenation.
 Lipid structures: 17 KB
Lipid structures: 17 KB
Triglycerides account for 98.3% of milkfat. The distribution of fatty acids on the triglyceride chain, while there are hundreds of different combinations, is not random. The fatty acid pattern is important when determining the physical properties of the lipids. In general, the SN1 position binds mostly longer carbon length fatty acids, and the SN3 position binds mostly shorter carbon length and unsaturated fatty acids. For example:
- C4 - 97% in SN3
- C6 - 84% in SN3
- C18 - 58% in SN1
The small amounts of mono- , diglycerides, and free fatty acids in fresh milk may be a product of early lipolysis or simply incomplete synthesis. Other classes of lipids include phospholipids (0.8%) which are mainly associated with the fat globule membrane, and cholesterol (0.3%) which is mostly located in the fat globule core.
The physical properties of milkfat can be summerized as follows:
- density at 20° C is 915 kg m(-3)*
- refractive index (589 nm) is 1.462 which decreases with increasing temperature
- solubility of water in fat is 0.14% (w/w) at 20° C and increases with increasing temperature
- thermal conductivity is about 0.17 J m(-1) s(-1) K(-1) at 20° C
- specific heat at 40° C is about 2.1kJ kg(-1) K(-1)
- electrical conductivity is <10(-12) ohm(-1) cm(-1)
- dielectric constant is about 3.1
*the brackets around numbers denote superscript
At room temperature, the lipids are solid, therefore, are correctly referred to as "fat" as opposed to "oil" which is liquid at room temperature. The melting points of individual triglycerides ranges from -75° C for tributyric glycerol to 72° C for tristearin. However, the final melting point of milkfat is at 37° C because higher melting triglycerides dissolve in the liquid fat. This temperature is significant because 37° C is the body temperature of the cow and the milk would need to be liquid at this temperature. The melting curves of milkfat are complicated by the diverse lipid composition:
- trans unsaturation increases melting points
- odd-numbered and branched chains decrease melting points
Crystallization of milkfat largely determines the physical stability of the fat globule and the consistency of high-fat dairy products, but crystal behaviour is also complicated by the wide range of different triglycerides. There are four forms that milkfat crystals can occur in; alpha, ß , ß ' 1, and ß ' 2, however, the alpha form is the least stable and is rarely observed in slowly cooled fat.
More than 95% of the total milk lipid is in the form of a globule ranging in size from 0.1 to 15 um in diameter. These liquid fat droplets are covered by a thin membrane, 8 to 10 nm in thickness, whose properties are completely different from both milkfat and plasma. The native fat globule membrane (FGM) is comprised of apical plasma membrane of the secretory cell which continually envelopes the lipid droplets as they pass into the lumen. The major components of the native FGM, therefore, is protein and phospholipids. The phospholipids are involved in the oxidation of milk. There may be some rearrangement of the membrane after release into the lumen as amphiphilic substances from the plasma adsorb onto the fat globule and parts of the membrane dissolve into either the globule core or the serum. The FGM decreases the lipid-serum interface to very low values, 1 to 2.5 mN/m, preventing the globules from immediate flocculation and coalescence, as well as protecting them from enzymatic action.
It is well known that if raw milk or cream is left to stand, it will separate. Stokes' Law predicts that fat globules will cream due to the differences in densities between the fat and plasma phases of milk. However, in cold raw milk, creaming takes place faster than is predicted from this fact alone. IgM, an immunoglobulin in milk, forms a complex with lipoproteins. This complex, known as cryoglobulin precipitates onto the fat globules and causes flocculation. This is known as cold agglutination. As fat globules cluster, the speed of rising increases and sweeps up the smaller globules with them. The cream layer forms very rapidly, within 20 to 30 min., in cold milk.
Homogenization of milk prevents this creaming by decreasing the diameter and size distribution of the fat globules, causing the speed of rise to be similar for the majority of globules. As well, homogenization causes the formation of a recombined membrane which is much similar in density to the continuous phase.
Recombined membranes are very different than native FGM. Processing steps such as homogenization, decreases the average diameter of fat globule and significantly increases the surface area. Some of the native FGM will remain adsorbed but there is no longer enough of it to cover all of the newly created surface area. Immediately after disruption of the fat globule, the surface tension raises to a high level of 15 mN/m and amphiphilic molecules in the plasma quickly adsorb to the lipid droplet to lower this value. The adsorbed layers consist mainly of serum proteins and casein micelles.
 Electron Micrograph of Homogenized Milkfat Globules 77 KB
Electron Micrograph of Homogenized Milkfat Globules 77 KB
While homogenization is the principal method for acheiving stabilization of the fat emulsion in milk, fat destabilization is necessary for structure formation inbutter, whipping cream and ice cream. Fat destabilization refers to the process of clustering and clumping (partial coalescence) of the fat globules which leads to the development of a continuous internal fat network or matrix structure in the product. Fat destabilization (sometimes "fat agglomeration") is a general term that describes the summation of several different phenomena. These include:
- Coalescence:
- an irreversible increase in the size of fat globules and a loss of identity of the coalescing globules;
- Flocculation:
- a reversible (with minor energy input) agglomeration/clustering of fat globules with no loss of identity of the globules in the floc; the fat globules that flocculate ; they can be easily redispersed if they are held together by weak forces, or they might be harder to redisperse to
they share part of their interfacial layers;
- Partial coalescence:
- an irreversible agglomeration/clustering of fat globules, held together by a combination of fat crystals and liquid fat, and a retention of identity of individual globules as long as the crystal structure is maintained (i.e., temperature dependent, once the crystals melt, the cluster coalesces). They usually come together in a shear field, as in whipping, and it is envisioned that the crystals at the surface of the droplets are responsible for causing colliding globules to stick together, while the liquid fat partially flows between they and acts as the "cement". Partial coalescence dominates structure formation in whipped, aerated dairy emulsions, and it should be emphasized that crystals within the emulsion droplets are responsible for its occurrence.
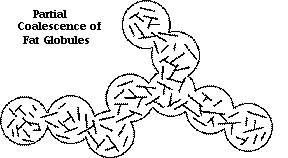
A good reference for more information on fat globules can be found in Mulder and Walstra.
Like all fats, milkfat provides lubrication. They impart a creamy mouth feel as opposed to a dry texture. Butter flavour is unique and is derived from low levels of short chain fatty acids. If too many short chain fatty acids are hydrolyzed (separated) from the triglycerides, however, the product will taste rancid. Butter fat also acts as a reservoir for other flavours, especially in aged cheese. Fat globules produce a 'shortening' effect in cheese by keeping the protein matrix extended to give a soft texture. Fat substitutes are designed to mimic the globular property of milk fat.
The spreadable range of butter fat is 16-24° C. Unfortunately butter is not spreadable at refrigeration temperatures. Milk fat provides energy (1g = 9 cal.), and nutrients (essential fatty acids, fat soluble vitamins).
Milk Proteins
The primary structure of proteins consists of a polypeptide chain of amino acids residues joined together by peptide linkages, which may also be cross-linked by disulphide bridges. Amino acids contain both a weakly basic amino group, and a weakly acid carboxyl group both connected to a hydrocarbon chain, which is unique to different amino acids. The three-dimensional organization of proteins, or conformation, also involves secondary, tertiary, and quaternary structures. The secondary structure refers to the spatial arrangement of amino acid residues that are near one another in the linear sequence. The alpha-helix and ß -pleated sheat are examples of secondary structures arising from regular and periodic steric relationships. The tertiary structure refers to the spatial arrangement of amino acid residues that are far apart in the linear sequence, giving rise to further coiling and folding. If the protein is tightly coiled and folded into a somewhat spherical shape, it is called a globular protein. If the protein consists of long polypeptide chains which are intermolecularly linked, they are called fibrous proteins. Quaternary structure occurs when proteins with two or more polypeptide chain subunits are associated.
Milk Protein Fractionation
The nitrogen content of milk is distributed among caseins (76%), whey proteins (18%), and non-protein nitrogen (NPN) (6%). This does not include the minor proteins that are associated with the FGM. This nitrogen distribution can be determined by the Rowland fractionation method:
- Precipitation at pH 4.6 - separates caseins from whey nitrogen
- Precipitation with sodium acetate and acetic acid (pH 5.0) - separates total proteins from whey NPN
The concentration of proteins in milk is as follows:
grams/ litre % of total protein
_______________________________________________________________________________
Total Protein 33 100
Total Caseins 26 79.5
alpha s1 10 30.6
alpha s2 2.6 8.0
beta 9.3 28.4
kappa 3.3 10.1
Total Whey Proteins 6.3 19.3
alpha lactalbumin 1.2 3.7
beta lactoglobulin 3.2 9.8
BSA 0.4 1.2
Immunoglobulins 0.7 2.1
Proteose peptone 0.8 2.4
_____________________________________________________________
Caseins, as well as their structural form - casein micelles, whey proteins, and milk enzymes will now be examined in further detail.
The casein content of milk represents about 80% of milk proteins. The principal casein fractions are alpha(s1) and alpha(s2)-caseins, ß -casein, and kappa-casein. The distinguishing property of all caseins is their low solubility at pH 4.6. The common compositional factor is that caseins are conjugated proteins, most with phosphate group(s) esterified to serine residues. These phosphate groups are important to the structure of the casein micelle. Calcium binding by the individual caseins is proportional to the phosphate content.
The conformation of caseins is much like that of denatured globular proteins. The high number of proline residues in caseins causes particular bending of the protein chain and inhibits the formation of close-packed, ordered secondary structures. Caseins contain no disulfide bonds. As well, the lack of tertiary structure accounts for the stability of caseins against heat denaturation because there is very little structure to unfold. Without a tertiary structure there is considerable exposure of hydrophobic residues. This results in strong association reactions of the caseins and renders them insoluble in water.
Within the group of caseins, there are several distinguishing features based on their charge distribution and sensitivity to calcium precipitation:
- alpha(s1)-casein: (molecular weight 23,000; 199 residues, 17 proline residues)
- Two hydrophobic regions, containing all the proline residues, separated by a polar region, which contains all but one of eight phosphate groups. It can be precipitated at very low levels of calcium.
- alpha(s2)-casein: (molecular weight 25,000; 207 residues, 10 prolines)
- Concentrated negative charges near N-terminus and positive charges near C-terminus. It can also be precipitated at very low levels of calcium.
- ß -casein: (molecular weight 24,000; 209 residues, 35 prolines)
- Highly charged N-terminal region and a hydrophobic C-terminal region. Very amphiphilic protein acts like a detergent molecule. Self association is temperature dependant; will form a large polymer at 20° C but not at 4° C. Less sensitive to calcium precipitation.
- kappa-casein: (molecular weight 19,000; 169 residues, 20 prolines)
- Very resistant to calcium precipitation, stabilizing other caseins. Rennet cleavage at the Phe105-Met106 bond eliminates the stabilizing ability, leaving a hydrophobic portion, para-kappa-casein, and a hydrophilic portion called kappa-casein glycomacropeptide (GMP), or more accurately, caseinomacropeptide (CMP).
Most, but not all, of the casein proteins exist in a colloidal particle known as the casein micelle. Its biological function is to carry large amounts of highly insoluble CaP to mammalian young in liquid form and to form a clot in the stomach for more efficient nutrition. Besides casein protein, calcium and phosphate, the micelle also contains citrate, minor ions, lipase and plasmin enzymes, and entrapped milk serum. These micelles are rather porous structures, occupying about 4 ml/g and 6-12% of the total volume fraction of milk.
The "casein sub-micelle" model has been prominent for the last several years, and is illustrated and described with the following link, but there is not universal acceptance of this model, and mounting research evidence to suggest that there is not a defined sub-micellar structure to the micelle at all. Another model of a more open structure is also defined with the following link.
In the submicelle model, it is thought that there are small aggregates of whole casein, containing 10 to 100 casein molecules, called submicelles. It is thought that there are two different kinds of submicelle; with and without kappa-casein. These submicelles contain a hydrophobic core and are covered by a hydrophilic coat which is at least partly comprised of the polar moieties of kappa-casein. The hydrophilic CMP of the kappa-casein exists as a flexible hair.
The open model also suggests there are more dense and less dense regions within the midelle, but there is less of a well-defined structure. In this model, calcium phosphate nanoclusters bind caseins and provide for the differences in density within the casein micelle.
 Casein Micelle Structure 17 KB
Casein Micelle Structure 17 KB
Colloidal calcium phosphate (CCP) acts as a cement between the hundreds or even thousands of submicelles that form the casein micelle. Binding may be covalent or electrostatic. Submicelles rich in kappa-casein occupy a surface position, whereas those with less are buried in the interior. The resulting hairy layer, at least 7 nm thick, acts to prohibit further aggregation of submicelles by steric repulsion. The casein micelles are not static; there are three dynamic equilibria between the micelle and its surroundings:
- the free casein molecules and submicelles
- the free submicelles and micelles
- the dissoved colloidal calcium and phosphate
The following factors must be considered when assessing the stability of the casein micelle:
- Role of Ca++:
- More than 90% of the calcium content of skim milk is associated in some way or another with the casein micelle. The removal of Ca++ leads to reversible dissociation of ß -casein without micellular disintegration. The addition of Ca++ leads to aggregation.
- H Bonding:
- Some occurs between the individual caseins in the micelle but not much because there is no secondary structure in casein proteins.
- Disulphide Bonds:
- alpha(s1) and ß-caseins do not have any cysteine residues. If any S-S bonds occur within the micelle, they are not the driving force for stabilization.
- Hydrophobic Interactions:
- Caseins are among the most hydrophobic proteins and there is some evidence to suggest they play a role in the stability of the micelle. It must be remembered that hydrophobic interactions are very temperature sensitive.
- Electrostatic Interactions:
- Some of the subunit interactions may be the result of ionic bonding, but the overall micellar structure is very loose and open.
- van der Waals Forces:
- No sucess in relating these forces to micellular stability.
- Steric stabilization:
- As already noted, the hairy layer interferes with interparticle approach.
There are several factors that will affect the stability of the casein micelle system:
- Salt content:
- affects the calcium activity in the serum and calcium phosphate content of the micelles.
- pH:
- lowering the pH leads to dissolution of calcium phosphate until, at the isoelectric point (pH 4.6), all phosphate is dissolved and the caseins precipitate.
- Temperature:
- at 4° C, beta-casein begins to dissociate from the micelle, at 0° C, there is no micellar aggregation; freezing produces a precipitate called cryo-casein.
- Heat Treatment:
- whey proteins become adsorbed, altering the behaviour of the micelle.
- Dehydration:
- by ethanol, for example, leads to aggregation of the micelles.
When two or more of these factors are applied together, the effect can also be additive.
Caseins are able to aggregate if the surface of the micelle is reactive. The Schmidt model further illustrates this.
![[image]](https://webarchive.library.unt.edu/eot2008/20090117061609im_/http://www.foodsci.uoguelph.ca/deicon/schmidt2.gif) Casein micelle aggregation 17 KB
Casein micelle aggregation 17 KB
Although the casein micelle is fairly stable, there are four major ways in which aggregation can be induced:
- chymosin - rennet or other proteolytic enzymes as in Cheese manufacturing
- acid
- heat
- age gelation
Chymosin, or rennet, is most often used for enzyme coagulation. During the primary stage, rennet cleaves the Phe(105)-Met(106) linkage of kappa-casein resulting in the formation of the soluble CMP which diffuses away from the micelle and para-kappa-casein, a distinctly hydrophobic peptide that remains on the micelle. The patch or reactive site, as illustrated in the above image, that is left on the micelles after enzymatic cleavage is necessary before aggregation of the paracasein micelles can begin.
During the secondary stage, the micelles aggregate. This is due to the loss of steric repulsion of the kappa-casein as well as the loss of electrostatic repulsion due to the decrease in pH. As the pH approaches its isoelectric point (pH 4.6), the caseins aggregate. The casein micelles also have a strong tendency to aggregate because of hydrophobic interactions. Calcium assists coagulation by creating isoelctric conditions and by acting as a bridge between micelles. The temperature at the time of coagulation is very important to both the primary and secondary stages. With an increase in temperature up to 40° C, the rate of the rennet reaction increases. During the secondary stage, increased temperatures increase the hydrophobic reaction. The tertiary stage of coagulation involves the rearrangement of micelles after a gel has formed. There is a loss of paracasein identity as the milk curd firms and syneresis begins.
Acidification causes the casein micelles to destabilize or aggregate by decreasing their electric charge to that of the isoelectric point. At the same time, the acidity of the medium increases the solubility of minerals so that organic calcium and phosphorus contained in the micelle gradually become soluble in the aqueous phase. Casein micelles disintegrate and casein precipitates. Aggregation occurs as a result of entropically driven hydrophobic interactions.
At temperatures above the boiling point casein micelles will irreversibly aggregate. On heating, the buffer capacity of milk salts change, carbon dioxide is released, organic acids are produced, and tricalcium phophate and casein phosphate may be precipitated with the release of hydrogen ions.
Age gelation is an aggregation phenomenon that affects shelf-stable, sterilized dairy products, such as concentrated milk and UHT milk products. After weeks to months storage of these products, there is a sudden sharp increase in viscosity accompanied by visible gelation and irreversible aggregation of the micelles into long chains forming a three-dimensional network. The actual cause and mechanism is not yet clear, however, some theories exist:
- Proteolytic breakdown of the casein: bacterial or native plasmin enzymes that are resistant to heat treatment may lead to the formation of a gel
- Chemical reactions: polymerization of casein and whey proteins due to Maillard type or other chemical reactions
- Formation of kappa-casein-ß -lactoglobulin complexes
An excellent source of information on casein micelle stability can be found in Walstra.
The proteins appearing in the supernatant of milk after precipitation at pH 4.6 are collectively called whey proteins. These globular proteins are more water soluble than caseins and are subject to heat denaturation. Native whey proteins have good gelling and whipping properties. Denaturation increases their water holding capacity. The principle fractions are ß -lactoglobulin, alpha-lactalbumin, bovine serum albumin (BSA), and immunoglobulins (Ig).
ß -Lactoglobulins: (MW - 18,000; 162 residues)
This group, including eight genetic variants, comprises approximately half the total whey proteins. ß -Lactoglobulin has two internal disulfide bonds and one free thiol group. The conformation includes considerable secondary structure and exists naturally as a noncovalent linked dimer. At the isoelectric point (pH 3.5 to 5.2), the dimers are further associated to octamers but at pH below 3.4, they are dissociated to monomers.
alpha-Lactalbumins: (MW - 14,000; 123 residues)
These proteins contain eight cysteine groups, all involved in internal disulfide bonds, and four tryptophan residues. alpha-Lactalbumin has a highly ordered secondary structure, and a compact, spherical tertiary structure. Thermal denaturation and pH <4.0 results in the release of bound calcium.
Enzymes are a group of proteins that have the ability to catalyze chemical reactions and the speed of such reactions. The action of enzymes is very specific. Milk contains both indigenous and exogenous enzymes. Exogenous enzymes mainly consist of heat-stable enzymes produced by psychrotrophic bacteria: lipases, and proteinases. There are many indigenous enzymes that have been isolated from milk. The most significant group are the hydrolases:
- lipoprotein lipase
- plasmin
- alkaline phosphatase
Lipoprotein lipase (LPL): A lipase enzyme splits fats into glycerol and free fatty acids. This enzyme is found mainly in the plasma in association with casein micelles. The milkfat is protected from its action by the FGM. If the FGM has been damaged, or if certain cofactors (blood serum lipoproteins) are present, the LPL is able to attack the lipoproteins of the FGM. Lipolysis may be caused in this way.
Plasmin: Plasmin is a proteolytic enzyme; it splits proteins. Plasmin attacks both ß -casein and alpha(s2)-casein. It is very heat stable and responsible for the development of bitterness in pasteurized milk and UHT processed milk. It may also play a role in the ripening and flavour development of certain cheeses, such as Swiss cheese.
Alkaline phosphatase: Phosphatase enzymes are able to split specific phosporic acid esters into phosphoric acid and the related alcohols. Unlike most milk enzymes, it has a pH and temperature optima differing from physiological values; pH of 9.8. The enzyme is destroyed by minimum pasteurization temperatures, therefore, a phosphatase test can be done to ensure proper pasteurization.
Lactose is a disaccharide (2 sugars) made up of glucose and galactose (which are both monosaccharides). Because of the anomeric carbon on the right side of the structure below, lactose can exist as two isomers, alpha, as shown, or beta, in which the hydroxyl on the anomeric carbon would point up on the ring structure shown below.
![[image]](https://webarchive.library.unt.edu/eot2008/20090117061609im_/http://www.foodsci.uoguelph.ca/deicon/Lactose.tiff)
It comprises 4.8 to 5.2% of milk, 52% of milk SNF, and 70% of whey solids. It is not as sweet as sucrose. When lactose is hydrolyzed by ß -D-galactosidase (lactase), an enzyme that splits these monosaccharides, the result is increased sweetness, and depressed freezing point.
One of its most important functions is its utilization as a fermentation substrate. Lactic acid bacteria produce lactic acid from lactose, which is the beginning of many fermented dairy products. Because of their ability to metabolize lactose, they have a competitive advantage over many pathogenic and spoilage organisms.
Some people suffer from lactose intolerance; they lack the lactase enzyme, hence they cannot digest lactose, or dairy products containing lactose.
Crystallization of lactose occurs in an alpha form which commonly takes a tomahawk shape. This results in the defect called sandiness. Lactose is relatively insoluble which is a problem in many dairy products, ice cream, sweetened condensed milk.
In addition to lactose, fresh milk contains other carbohydrates in small amounts, including glucose, galactose, and oligosaccharides.
Vitamins are organic substances essential for many life processes. Milk includes fat soluble vitamins A , D, E, and K. Vitamin A is derived from retinol and ß -carotene. Because milk is an important source of dietary vitamin A, fat reduced products which have lost vitamin A with the fat, are required to supplement the product with vitamin A.
Milk is also an important source of dietary water soluble vitamins:
- B1 - thiamine
- B2 - riboflavin
- B6 - pyridoxine
- B12 - cyanocobalamin
- niacin
- pantothenic acid
There is also a small amount of vitamin C (ascorbic acid) present in raw milk but is very heat-labile and easily destroyed by pasteurization.
The vitamin content of fresh milk is given below:
Vitamin Contents per litre
---------------------------------------------------------------------------------
A (ug RE) 400
D (IU) 40
E (ug) 1000
K (ug) 50
B1 (ug) 450
B2 (ug) 1750
Niacin (ug) 900
B6 (ug) 500
Pantothenic acid (ug) 3500
Biotin (ug) 35
Folic acid (ug) 55
B12 (ug) 4.5
C (mg) 20
All 22 minerals considered to be essential to the human diet are present in milk. These include three families of salts:
- Sodium (Na), Potassium (K) and Chloride (Cl):These free ions are negatively correlated to lactose to maintain osmotic equilibrium of milk with blood.
- Calcium (Ca), Magnesium (Mg), Inorganic Phosphorous (P(i)), and Citrate: This group consists of 2/3 of the Ca, 1/3 of the Mg, 1/2 of the P(i), and less than 1/10 of the citrate in colloidal (nondiffusible) form and present in the casein micelle.
- Diffusible salts of Ca, Mg, citrate, and phosphate: These salts are very pH dependent and contribute to the overall acid-base equilibrium of milk.
The mineral content of fresh milk is given below:
Mineral Content per litre
---------------------------------------------------------------------------------
Sodium (mg) 350-900
Potassium (mg) 1100-1700
Chloride (mg) 900-1100
Calcium (mg) 1100-1300
Magnesium (mg) 90-140
Phosphorus (mg) 900-1000
Iron (ug) 300-600
Zinc (ug) 2000-6000
Copper (ug) 100-600
Manganese (ug) 20-50
Iodine (ug) 260
Fluoride (ug) 30-220
Selenium (ug) 5-67
Cobalt (ug) 0.5-1.3
Chromium (ug) 8-13
Molybdenum (ug) 18-120
Nickel (ug) 0-50
Silicon (ug) 750-7000
Vanadium (ug) tr-310
Tin (ug) 40-500
Arsenic (ug) 20-60
Physical Properties
The density of milk and milk products is used for the following;
- to convert volume into mass and vice versa
- to estimate the solids content
- to calculate other physical properties (e.g. kinematic viscosity)
Density, the mass of a certain quantity of material divided by its volume, is dependant on the following:
- temperature at the time of measurement
- temperature history of the material
- composition of the material (especially the fat content)
- inclusion of air (a complication with more viscous products)
With all of this in mind, the density of milk varies within the range of 1027 to 1033 kg m(-3) at 20° C.
The following table gives the density of various fluid dairy products as a function of fat and solids-not-fat (SNF) composition:
Product Density (kg/L) at:
Composition _______________________________
Product Fat (%) SNF (%) 4.4° C 10° C 20° C 38.9° C
_________________________________________________________________________________
Producer milk 4.00 8.95 1.035 1.033 1.030 1.023
Homogenized milk 3.6 8.6 1.033 1.032 1.029 1.022
Skim milk, pkg 0.02 8.9 1.036 1.035 1.033 1.026
Fortified skim 0.02 10.15 1.041 1.040 1.038 1.031
Half and half 12.25 7.75 1.027 1.025 1.020 1,010
Half and half, fort. 11.30 8.9 1.031 1.030 1.024 1.014
Light cream 20.00 7.2 1.021 1.018 1.012 1.000
Heavy cream 36.60 5.55 1.008 1.005 0.994 0.978
_________________________________________________________________________________
Viscosity of milk and milk products is important in determining the following:
- the rate of creaming
- rates of mass and heat transfer
- the flow conditions in dairy processes
Milk and skim milk, excepting cooled raw milk, exhibit Newtonian behavior, in which the viscosity is independent of the rate of shear. The viscosity of these products depends on the following:
- Temperature:
- cooler temperatures increase viscosity due to the increased voluminosity of casein micelles
- temperatures above 65° C increase viscosity due to the denaturation of whey proteins
- pH: an increase or decrease in pH of milk also causes an increase in casein micelle voluminosity
Cooled raw milk and cream exhibit non-Newtonian behavior in which the viscosity is dependant on the shear rate. Agitation may cause partial coalescence of the fat globules (partial churning) which increases viscocity. Fat globules that have under gone cold agglutination, may be dispersed due to agitation, causing a decrease in viscosity.
Freezing point depression is a colligative property which is determined by the molarity of solutes rather than by the percentage by weight or volume. In the dairy industry, freezing point of milk is mainly used to determine added water but it can also been used to determine lactose content in milk, estimate whey powder contents in skim milk powder, and to determine water activity of cheese.
The freezing point of milk is usually in the range of -0.512 to -0.550° C with an average of about -0.522° C.
Correct interpretation of freezing point data with respect to added water depends on a good understanding of the factors affecting freezing point depression. With respect to interpretation of freezing points for added water determination, the most significant variables are the nutritional status of the herd and the access to water. Under feeding causes increased freezing points. Large temporary increases in freezing point occur after consumption of large amounts of water because milk is iso-osmotic with blood.
The primary sources of non-intentional added water in milk are residual rinse water and condensation in the milking system.
Both titratable acidity and pH are used to measure milk acidity. The pH of milk at 25° C normally varies within a relatively narrow range of 6.5 to 6.7. The normal range for titratable acidity of herd milks is 13 to 20 mmol/L. Because of the large inherent variation, the measure of titratable acidity has little practical value except to measure changes in acidity (eg., during lactic fermentation) and even for this purpose, pH is a better measurement.
There are many components in milk which provide a buffering action. The major buffering groups of milk are caseins and phosphate.
Optical properties provide the basis for many rapid, indirect methods of analysis such as proximate analysis by infrared absorbency or light scattering. Optical properties also determine the appearance of milk and milk products. Light scattering by fat globules and casein micelles causes milk to appear turbid and opaque. Light scattering occurs when the wave length of light is near the same magnitude as the particle. Thus, smaller particles scatter light of shorter wavelengths. Skim milk appears slightly blue because casein micelles scatter the shorter wavelengths of visible light (blue) more than the red. The carotenoid precursor of vitamin A, ß -carotene, contained in milk fat, is responsible for the 'creamy' colour of milk. Riboflavin imparts a greenish colour to whey.
Refractive index (RI) is normally determined at 20° C with the D line of the sodium spectrum. The refractive index of milk is 1.3440 to 1.3485 and can be used to estimate total solids.
More information on dairy chemistry can be found in Walstra and Jenness, Goff and Hill, and Wong et al.
Back to the Home Page of ![[Dairy Home Page]](https://webarchive.library.unt.edu/eot2008/20090117061609im_/http://www.foodsci.uoguelph.ca/deicon/dairyicon3.gif)
{kind=link}