
| | | | |
Research
|
| New Insight into Intrachromosomal Deletions Induced by Chrysotile in the gpt delta Transgenic Mutation Assay An Xu,1,2 Lubomir B. Smilenov,1 Peng He,1 Ken-ichi Masumura,3 Takehiko Nohmi,3 Zengliang Yu,2 and Tom K. Hei1,4 1Center for Radiological Research, College of Physicians & Surgeons, Columbia University, New York, New York, USA; 2Key Laboratory of Ion Beam Bioengineering, Institute of Plasma Physics, Chinese Academy of Sciences, Hefei, Anhui, People's Republic of China; 3Division of Genetics and Mutagenesis, National Institute of Health Sciences, Tokyo, Japan; 4Department of Environmental Health Sciences, Mailman School of Public Health, Columbia University, New York, New York, USA Abstract
Background: Genotoxicity is often a prerequisite to the development of malignancy. Considerable evidence has shown that exposure to asbestos fibers results in the generation of chromosomal aberrations and multilocus mutations using various in vitro approaches. However, there is less evidence to demonstrate the contribution of deletions to the mutagenicity of asbestos fibers in vivo. Objectives: In the present study, we investigated the mutant fractions and the patterns induced by chrysotile fibers in gpt delta transgenic mouse primary embryo fibroblasts (MEFs) and compared the results obtained with hydrogen peroxide (H2O2) in an attempt to illustrate the role of oxyradicals in fiber mutagenesis. Results: Chrysotile fibers induced a dose-dependent increase in mutation yield at the redBA/gam loci in transgenic MEF cells. The number of 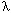 mutants losing both redBA and gam loci induced by chrysotiles at a dose of 1 µg/cm2 increased by > 5-fold relative to nontreated controls (p < 0.005) . Mutation spectra analyses showed that the ratio of mutants losing the redBA/gam region induced by chrysotiles was similar to those induced by equitoxic doses of H2O2. Moreover, treatment with catalase abrogated the accumulation of mutants losing both redBA and gam loci induced by chrysotiles at a dose of 1 µg/cm2 increased by > 5-fold relative to nontreated controls (p < 0.005) . Mutation spectra analyses showed that the ratio of mutants losing the redBA/gam region induced by chrysotiles was similar to those induced by equitoxic doses of H2O2. Moreover, treatment with catalase abrogated the accumulation of  -H2AX, a biomarker of DNA double-strand breaks, induced by chrysotile fibers. -H2AX, a biomarker of DNA double-strand breaks, induced by chrysotile fibers. Conclusions: Our results provide novel information on the frequencies and types of mutations induced by asbestos fibers in the gpt delta transgenic mouse mutagenic assay, which shows great promise for evaluating fiber/particle mutagenicity in vivo. Key words: chrysotile asbestos, gpt delta transgenic mutation system, kilobase-sized mutation, oxyradicals, -H2AX. Environ Health Perspect 115:87–92 (2007) . doi:10.1289/ehp.9425 available via http://dx.doi.org/ [Online 6 September 2006]
Address correspondence to T.K. Hei, Center for Radiological Research, Columbia University, New York, NY 10032 USA. Telephone: (212) 305-8462. Fax: (212) 305-3229. E-mail: TKH1@Columbia.edu We thank M. Partridge and V. Ivanov for their critical reading of the manuscript and R. Baker for his technical assistance with Spi– detection. Work was supported in part by National Institutes of Health grant ES 05786, Superfund grant ES 10349, National Institute of Environmental Health Sciences Center grant ES 09089, and National Nature Science Foundation of China grant 20322202. The authors declare they have no competing financial interests. Received 14 June 2006 ; accepted 6 September 2006. |
|
|
|
Asbestos fibers, a group of naturally occurring silicate minerals, are well-established human carcinogens. They are causally related to the development of asbestosis, bronchial carcinoma, malignant mesothelioma of the pleura and peritoneum, and possibly cancers of the gastrointestinal tract and larynx (Gustavsson et al. 1998; International Agency for Research on Cancer 1987). Although the main concern of asbestos-related diseases focuses primarily on the workplace, the danger of developing such diseases now extends beyond that of a simple occupational hazard because accumulating evidence suggests that asbestos fibers are widely distributed in the environment to which the general public may be exposed (Gardner and Saracci 1989). Individuals may be subjected to prolonged exposure to asbestos in their homes, schools, drinking water, neighborhoods of industrial sources of asbestos, or areas of a natural occurring asbestos (Kjaerheim et al. 2005; Miller 2005; Rom et al. 2001). Recent evidence indicates an excess risk of mesothelioma in individuals living in the vicinity of a natural occurring source of asbestos (Pan et al. 2005). The continued discovery of routes through which the general public may be exposed to asbestos suggests a long-term, low-level exposure of a large number of people, which may lead to an elevated risk of asbestos-related diseases.
The mechanisms by which asbestos produces malignancy are unclear at present. It has been reported that fiber dimension, biopersistence, composition, surface reactivity, and physical durability are important criteria for the carcinogenicity of the fibers, indicating that carcinogenic mechanisms of asbestos are likely to be complex and involve multiple pathways (Bernstein et al. 2003; Coin et al. 1994). Various highly quantitative genotoxicity assays ranging from DNA strand breaks to gene mutations in rodent cells have been performed to estimate the carcinogenic potential of asbestos fibers (Lezon-Geyda et al. 1996; Poser et al. 2004). Although asbestos fibers have been shown to induce chromosomal aberrations and sister chromatid exchanges, mutagenic studies at the hypoxanthine–guanine phosphoribosyl transferase (hprt; GenBank accession no. NM_013556; http://www.ncbi.nlm.nih.gov/) and ouabain loci in mammalian cells have been shown to be either inactive or weakly active (Jaurand 1996). Using the human–hamster hybrid cell line (AL) in which mutations are scored at a marker gene (CD59) located on human chromosome 11 (11p13) that the AL cell carries as its only human chromosome, our previous studies have demonstrated that asbestos fibers are indeed mutagenic and induce mostly deletions involving millions of base pairs (Hei et al. 1992). In contrast, among the same fiber-treated AL cell population, there are few, if any, mutations scored at the hprt locus of the hamster X chromosome. This discrepancy has been attributed to the possibility that the hprt gene is located on the X chromosome, and large deletions in the region of the gene that are required for cell survival would be lethal and any mutants induced would not be viable. In recent years several other mutagenic assays that are proficient in detecting either large deletions, homologous recombinations, or score mutants located in nonessential genes have been used successfully to demonstrate the mutagenic potential of various fiber types (Lezon-Geyda et al. 1996; Park and Aust 1998). These findings provide a direct link between chromosomal abnormalities that frequently have been demonstrated in fiber exposed human and rodent cell lines and carcinogenicity in vivo. However, there is less direct evidence that illustrates chromosomal mutations of asbestos fibers in various organs and tissues in intact organisms.
The use of transgenic mouse systems carrying bacterial reporter genes such as lacZ, lacI, and cII has opened a promising opportunity for short-term mutagenicity analysis (Dean et al. 1999). There is evidence to show that asbestos fibers are mutagenic and induce point mutations in either Big Blue transgenic mice or rats bearing -lacI as a reporter gene (Rihn et al. 2000; Topinka et al. 2004; Unfried et al. 2002). However, most genomewide mutations such as large deletions, insertions, translocations, and aneuploidy are not effectively recovered by the lacI shuttle vector. To efficiently recover large deletions in vivo, gpt (xanthine phosphoribosyltransferase; GenBank accession no. NP_414773; http://www.ncbi.nlm.nih.gov/GenBank) delta transgenic mice have been established by integrating multiple copies of EG10 DNA with the redBA and gam (Genbank accession no. J02459; http://www.ncbi.nlm.nih.gov/GenBank) genes into each chromosome 17 of C57BL/6J mice (Nohmi and Masumura 2004). Because wild-type -phage DNA replicate poorly in the presence of P2 lysogens in the host cells (called "sensitive to P2 interference" or "Spi"), only mutant phages that are deficient in the functions of both the redBA and gam genes are able to escape from P2 interference (called "Spi–") and form visible clear plaques on a bacterial lawn. Simultaneous inactivations of both the redBA and gam genes, an indication of deletions in the gene loci region, provide an available method to quantify deletion mutations induced by various physical and chemical mutagens, such as X rays and alkylating agents (Horiguchi et al. 2001; Shibata et al. 2005).
Chrysotile asbestos, a fibrous serpentine, is the most commercially used form of asbestos in the world trade and accounts for > 95% of asbestos found in United States buildings. In the present study we adapted the gpt delta transgenic mouse mutation system to evaluate the genotoxicity of chrysotile in gpt delta mouse primary embryo fibroblast (MEF) cells. We investigated the mutation frequencies at both redBA and gam (GenBank accession no. J02459; http://www.ncbi.nlm.nih.gov/GenBank) loci and the contribution of deletions > 2 kb to the mutagenicity of chrysotile fibers. Because reactive oxygen species (ROS) such as superoxide anions (O2–) and hydrogen peroxide (H2O2) originate not only from redox reactions catalyzed on the fiber surface but also from the incomplete phagocytosis of fibers in various cells, such as phagocytic, mesothelial, and rat lung epithelium cells, we speculated that asbestos fibers would induce similar types of mutations as that of chemically generated oxyradicals. We found that both chrysotile and H2O2 dramatically increased the mutation yield, which could be abrogated by concurrent treatment with catalase. Furthermore, the ratios of mutants with deletions > 2 kb were similar to those generated by oxyradicals at two equitoxic doses. The accumulation of phosphorylated histone H2AX (-H2AX) further demonstrated the involvement of DNA double-strand break (DSB) in the mutagenicity of chrysotiles. These results provide direct evidence that asbestos fibers induced kilobase pair deletion mutations in a transgenic mouse mutation system, and that these were mediated by oxyradicals.
MEF cell culture. gpt delta transgenic mice, obtained from T. Nohmi, were mated, and pregnant females were sacrificed on day 14 of the experimental protocol were previously approved by the Columbia University Institutional Animal Care and Use Committee. The animals were treated humanely and with regard for the alleviation of pain and suffering. The embryos were surgically removed and embryonic tissue prepared in culture according to standard procedures (Hogan et al. 1994). These cultures were grown and maintained in Dulbecco's modified Eagle's medium (Gibco-BRL, Gaithersburg, MD, USA) containing 15% heat-inactivated fetal bovine serum and penicillin (100 U/mL), streptomycin (50 µg/mL) in a 5% CO2 environment at 37°C.
Chrysotile preparation. We used International Union Against Cancer standard reference chrysotile asbestos (average length, 7.8 µm; average diameter, 0.2 µm) in these studies (Timbrell 1979). The fibers were prepared as described previously (Hei et al. 1992). Briefly, samples of fibers were weighed and suspended in distilled water. The fiber suspension was triturated 6–8 times with a 20-gauge syringe needle. A stock solution of the fibers was sterilized by autoclaving and mixed to ensure a uniform suspension before being diluted with tissue culture medium for cell treatment.
Cytotoxicity assay. We evaluated cell viability using MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay on the basis of the ability of viable cells to convert a water-soluble tetrazolium salt into a water-insoluble formazan product (Scudiero et al. 1988). The enzymatic reduction of the tetrazolium salt happens only in living, metabolically active cells but not in dead cells. Cultures were incubated in two-well chamber slides at a density of 1.0  105 cells per well at 37°C for 24 hr. Graded doses of either chrysotile or H2O2 were added to the culture medium and incubated for another 24 hr in the presence of serum (chrysotile) or 15 min in the absence of serum (H2O2). At the end of the treatment period, the medium was removed, 200 µL of 5 mg/mL MTT was added to each well, and the cultures were incubated for another 4 hr. The supernatant was removed and 1 mL acidic isopropanol was added to dissolve the formazan crystals. The absorbance at 570 nm was determined by an Ultrospec 3100 pro UV/Visible spectrophotometer (Biochrom Ltd., Cambridge, UK). 105 cells per well at 37°C for 24 hr. Graded doses of either chrysotile or H2O2 were added to the culture medium and incubated for another 24 hr in the presence of serum (chrysotile) or 15 min in the absence of serum (H2O2). At the end of the treatment period, the medium was removed, 200 µL of 5 mg/mL MTT was added to each well, and the cultures were incubated for another 4 hr. The supernatant was removed and 1 mL acidic isopropanol was added to dissolve the formazan crystals. The absorbance at 570 nm was determined by an Ultrospec 3100 pro UV/Visible spectrophotometer (Biochrom Ltd., Cambridge, UK).
Genomic DNA isolation. We isolated genomic DNA from MEF cells using the RecoverEase DNA isolation kit (Stratagene, La Jolla, CA, USA) according to the protocol developed by the supplier. Briefly, about 5.0 106 cells were transferred to a chilled Wheaton dounce tissue grinder (Fisher, Hampton, NH, USA), and the homogenate obtained was filtered and centrifuged at 1,100 g for 12 min at 4°C. The pellet was resuspended in digestion buffer containing RNAses (RANse-It; Stratagene) containing proteinase K solution (2 mg/mL prewarmed to 50°C). Using wide-bore pipette tips, the samples were transferred to dialysis cups floating on the surface of TE buffer (500 mL) and dialyzed for 24 hr. The purity and concentration of DNA was checked spectrophotometrically and samples were diluted with TE [10 mM Tris–Cl (pH 7.5), 1 mM EDTA] buffer to a final DNA concentration of approximately 0.5 mg/mL, and stored at 4°C for up to 3 months prior to mutation analysis.
In vitro packaging of DNA. The DNA was recovered from approximately 5 µg genomic DNA and packaged with terminase and phage proteins contained in the Transpack kit (Stratagene) to produce infectious phages. Viable phages were infected into Escherichia coli XL-1 Blue MRA (Stratagene), mixed with -trypticase agarose and poured onto 100-mm plates containing 30 mL bottom agar. Plates were incubated overnight at 37°C. The average of rescued phages per packaging reaction was 1.8 106 in the present studies. There was no significant difference in the titers between control and exposed groups.
Spi– mutation analysis. The mutant frequencies at red/gam loci were determined by Spi– selection as described previously (Nohmi and Masumura 2004; Nohmi et al. 1996; Shibata et al. 2005). Briefly, packaged phages were infected into E. coli XL-1 Blue MRA (P2) (Stratagene). Infected cells were mixed with molten soft agar, poured onto -trypticase agar plates and incubated at 37°C. The plaques detected on the plates (Spi– candidates) were suspended in 50 µL of SM buffer [0.58% NaCl, 0.2% MgSO4•7H2O, 50 mM Tris–HCl, 0.01% gelatin (pH 7.5)]. The suspension was spotted on the two types of plates where E. coli XL-1 Blue MRA (P2) or WL95 (P2) strain was spread. The plates were incubated for 24 hr at 37°C. The numbers of mutants that made clear spots on both strains were counted as confirmed Spi– mutants. Mutation frequencies were calculated by comparing the titration and number of confirmed mutant plaques.
Spi– mutant characterization. To determine the mutated region, the phage DNA was used and subjected to DNA sequence and polymerase chain reaction (PCR) analysis with various sets of primers (Horiguchi et al. 2001). The PCR primers used were as follows: primer 1: 5´-CACTCTCTTTGATGCGAATGCCAGCGTCAGAC-3´; primer 2: 5´-CAGGAGTAATTATGCGGAACAGAATCATGCCTGGTG-3´; primer 3: 5´-GTGAGGATGCGTCATCGCCATTGCTCCCC-3´; primer 4: 5´-GCGATGAAAAGATGTTTCGTGAAGCCGTCGACGC-3´; primer 5: 5´-AAACAGGCGCGGGCATCAGCGTGGTCTGA-39´; primer 12: 5´-CGCGCGGTCGAGGGACCTAATAACTTCGTA-3´. We performed PCR amplification under the following conditions: 4 µL of phage DNA, 0.2 mM each dNTP, 1.5 mM MgCl2, Taq DNA polymerase (or ExTaq; Takara Shuzo Co., Kyoto, Japan), and 20 pmol of each primer in a 40-µL reaction volume; heating for 1–2 min at 94°C, and 24 cycles at 98°C for 20 sec and at 68°C for X minutes (1 min/1 kb), followed by final extension at 72°C for 10 min. The products were analyzed using agarose gel electrophoresis.
The PCR products were sequenced by ABI's 3100 capillary sequencers (Dye Terminator Cycle Sequencing; PE Applied Biosystems, Foster City, CA, USA). PCR products for templates of sequence were purified using PCR product presequencing kit (Amersham Life Science, Piscataway, NJ, USA).
Sequence primers are as follows: s102: 5´-AATCCAAACTCTTTACCCGTCCTTGGGT-3´; s201: 5´-CGCTTGATAACTCT
GTTGAATGGCTCT-3´; s301: 5´-GGTGGAATCCCATCAGCGTTACCGTTT-3´; s302: 5´-AGTGATTGCGCCTACCCGGATATTATCGTG-3´; s403: 5´-CCAGCCGACACGTTCAGCCAGCTTCCCAG-3´.
The entire DNA sequence of EG10 is available at http://dgm2alpha.nihs.go.jp (Masumura et al. 2003).
Determination of H2AX phosphorylation using flow cytometry. The cells were fixed by adding 2% paraformaldehyde dropwise while being vortexed. The fixed cells were then stained with mouse monoclonal anti–-H2AX (Upstate, Lake Placid, NY, USA) and fluorescein isothiocyanate (FITC)–conjugated secondary antibodies (Sigma-Aldrich Chemical Co., St. Louis, MO, USA) as described by Kurose et al. (2005). The cells were then suspended in 0.5 mL of 10 µg/mL propidium iodide (PI) and 40 µg/mL RNase A and incubated at 4°C for at least 30 min. The fluorescence of PI and FITC of individual cells induced by excitation with a 488-nm argon ion laser was measured using a FACSCalibur cytometer (BD Biosciences, San Jose, CA).
Statistical analysis. All numerical data were calculated as mean and SD and evaluated by Student's t-test. The statistical significance was tested at p < 0.05 as the critical value.
Chrysotile-induced dose-dependent toxicity in transgenic MEF cells. The viability of MEF cells exposed to graded doses of chrysotile was analyzed by using the MTT assay. As shown in Figure 1, exposure of MEF cells to doses of chrysotiles ranging from 0.5 to 6 µg/cm2 for 24 hr produced a dose-dependent decrease in cell viability. The viabilities of MEF cells was reduced by 14, 29, and 59%, when the concentrations of chrysotile were 0.5, 1, and 2 µg/cm2, respectively. The median lethal dose of chrysotile, which resulted in 50% cell killing, was approximately 3.2 µg/cm2.
|

Figure 1. Cell viability of transgenic MEF cells treated with graded doses of chrysotile for 24 hr. Data were the average of three independent experiments. Error bars indicate ± SD.
|
|

Figure 2. Mutagenic potential of chrysotile asbestos at redBA and gam loci in transgenic MEF cells. MEF cells, 5 105, were treated with graded doses of chrysotile as described in the text. Results were expressed as the total number of confirmed mutants divided by the total number of rescued phages. The average number of preexisting mutants per 106 plaques used for these experiments was 4.69 ± 1.42. Data were pooled from six independent experiments. Error bars indicate ± SD.
*Significantly different at p < 0.05.
|
Mutation frequencies at red/gam gene loci were elevated in response to chrysotile exposure. We have shown previously that asbestos is mutagenic and induces multilocus deletions in mammalian cells (Hei et al. 1992). To investigate the mutagenicity of asbestos in the gpt delta assay, we used an Spi– mutation assay to determine the mutation frequencies induced by chrysotile exposure in transgenic MEF cells. The average number of spontaneous red/gam gene mutants per 106 recovered plaques in MEF cells used for these experiments was 4.69 ± 1.80. Treatment of MEF cells with chrysotile fibers resulted in a dose-dependent induction of mutation yield atthe red/gam gene locus (Figure 2). A significant increase in mutation yield over the background level was observed at fiber concentrations > 1 µg/cm2 (p < 0.005). The mutant fraction in cells treated with a dose of 1 µg/cm2 of fibers was 2.4-fold higher than background. These results indicated that chrysotile asbestos were able to produce deletion mutations in gpt delta transgenic mutation assay system.
Characterization of mutant spectra induced by chrysotile. To determine the spectrum of mutations induced by chrysotile fibers, 93 and 74 mutants from control cells and cells treated with chrysotile at 1 µg/cm2, respectively, were subjected to either PCR analysis or DNA sequence analysis. The PCR product of redBA/gam in the wild-type EG10 was approximately 2 kb. If a PCR product did not show any discrete alteration on the gel, the mutant was classified as one containing a point mutation with either a base substitution or a frameshift causing no alteration in the size of the gene product. In contrast an absence of visible PCR product was taken as evidence of a mutant with a deletion > 2 kb as a result of losing both redBA and gam genes. The types of mutations identified from analysis of these mutants are listed in Table 1 and Figure 3. To minimize the possibility that these isolated mutants were spontaneously derived, we selected mutant phages from only the dose of chrysotile that resulted in the highest inductions over background levels. The majority of spontaneous mutants were deletions of various sizes throughout the red BA/gam genes (86 of 93 or 92%). Of these deletion mutants, 1 bp deletion made up 68 of 93 or 73%, whereas deletions ranging from 2 bp to 1 kb made up 8 of 93 or 8.6%. Of the spontaneous mutations with deletions 10 of 93 or 11% encompass regions of both the gam and redBA genes. In contrast, 41 of 74 or 56% and 10 of 74 or 14% of mutants recovered from chrysotile treated cells were single base pair deletion and deletions ranging from 2 bp to 1 kb, respectively. The proportion of mutants induced by chrysotile suffering loss of both the gam and redBA genes was increased from 10 of 93 or 11% among spontaneous mutants to 17 of 74 or 23% in fiber-treated MEF cells (Table 1).
Table 1.

|
|

Figure 3. Schematic map of EG10 transgene. Abbreviations: bio, genetic marker used in bacteriophage lambda vectors; CAT, chloramphenicol acetyltransferase (GenBank accession no. AJ401050; http://www.ncbi.nlm.nih.gov/GenBank/); cro, transcription inhibitor; gpt, xanthine phosphoribosyltransferase (GenBank accession no. NP_414773); J, codes for phage tail gene; loxP, locus of X over P1, a site on the bacteriophage P1 consisting of 34 bp; redA, redB, and gam, single copy bacteriophage genes.
|
Table 2.

|
|

Figure 4. Mutant fractions at redBA/gam loci in MEF cells exposed to either chrysotile asbestos at a dose of 1 µg/cm2 or 2.9 mM H2O2 either in the presence (+) or absence (–) of catalase (5,000 U/mL). Results were expressed as the total number of confirmed -phage mutants divided by the total number of rescued phages. The average number of preexisting mutants per 106 plaques used for these experiments was 4.69 ± 1.42. Numbers above error bars indicate the percentage of relative viability. Data were pooled from three to six independent experiments. Error bars indicate ± SD.
|
|

Figure 5. Mutant fractions of -phage mutants with specific molecular characteristics in MEF cells exposed to either chrysotile at a concentration of 1 µg/cm2 or H2O2 at a dose of 2.9 mM. Numbers above error bars indicate ratio of mutation type calculated as percentage. Data were pooled from three to six independent experiments. Error bars indicate ± SD.
|
|

Figure 6. Accumulation of -H2AX in MEF cells exposed to either chrysotile for 24 hr in completed medium or H2O2 for 15 min in serum-free medium in the presence or absence of catalase. (A) Generation of -H2AX in cells treated with either chrysotile or H2O2 detected by flow cytometry using mouse monoclonal anti–-H2AX labeled with FITC. Results are shown using FL1-H. (B) Induced -H2AX in MEF cells exposed to either chrysotile or H2O2 in the presence or absence of catalase. (1) Control; (2) 1 µg/cm2 chrysotile; (3) 2 µg/cm2 chrysotile; (4) 5,000 U/mL catalase; (5) 1 µg/cm2 chrysotile + 5,000 U/mL catalase; (6) 29 mM hydrogen peroxide; (7) 29 mM hydrogen peroxide + 5,000 U/mL catalase. Data were pooled from three independent experiments. Error bars indicate ± SD.
|
Deletions > 2 kb contribute to chrysotile-induced mutagenicity. To provide further evidence of the contribution of deletions > 2 kb to the mutagenicity of chrysotile, we compared the frequencies of deletions > 2 kb induced by chrysotile at a dose of 1 µg/cm2 with those derived spontaneously from control cultures (Table 2). Although the total Spi– mutant yield in chrysotile-treated cells was 2.4-fold higher than that of controls, the frequency of deletions > 2 kb induced by 1 µg/cm2 of fibers was 5.2-fold higher than those derived from nontreated control (2.6 vs. 0.5 10–6, p < 0.005). The frequency of base substitution and small deletions including single base deletions and deletions < 1 kb formed in fiber-treated MEF cells was only 2-fold higher than those from nontreated cases. These results indicated that the major types of mutations induced by chrysotile were deletions > 2 kb.
Oxyradicals mediated the mutagenicity of chrysotile in transgenic mouse mutation assay system. There is evidence that the genotoxicity/ carcinogenicity of asbestos fibers is mediated by reactive oxygen/nitrogen species (Shukla et al. 2003). To demonstrate that oxyradicals mediated the mutagenicity of chrysotile fibers in MEF cells, we exposed MEF cells to either chrysotile for 24 hr in complete medium, or to H2O2 in serum free medium for 15 min in the presence or absence of catalase (Figure 4). The relative viability of MEF cells treated with a 1 µg/cm2 dose of chrysotile was 71%, whereas the relative viability of MEF cells after exposing to 2.9 mM H2O2 was 69%. Both chrysotile and H2O2 led to significant increases in Spi– mutant yields in MEF cells. As shown in Figure 4, the mutation yield induced by H2O2 treatment was slightly higher than that of chrysotile at equal toxic doses, although the difference was not statistically significant. Furthermore, the mutation yields induced by either chrysotile at a dose of 1 µg/cm2 or 2.9 mM H2O2 were dramatically suppressed in the presence of 5,000 U/mL catalase (p < 0.05). Interestingly, the ratio of the mutants with deletions > 2 kb was similar between chrysotile and H2O2 in that 20 of 84 or 24% of the mutants induced by 2.9 mM H2O2 lost both redBA and gam genes compared with 17 of 74 or 23% among those induced by a 1-µg/cm2 dose of chrysotile (Figure 5). The mutant fractions with deletions > 2 kb increased from 0.5 ± 0.16 observed in controls to either 2.6 ± 0.93 or 3.2 ± 1.34 in cells treated with either chrysotile or H2O2, respectively. The dose of catalase used here had little effect on the level of cell viability and mutant fraction in control cells. Similarly, heat-inactivated catalase (by boiling for 10 min) had little effect on the mutant fraction in exposed cells.
Induction of -H2AX in MEF cells. Among various type of DNA damages, the DSBs in DNA may be the most damaging and genotoxic, which elevate the frequencies of gene translocations, rearrangements, amplifications, and deletions during repair and misrepair of DSBs (Khanna and Jackson 2001). A very early step in the response of mammalian cells to DNA DSBs is the phosphorylation of histone H2AX at serine-139 at the sites of DNA damage. To investigate whether chrysotile induces phosphorylation of H2AX in MEF cells, we exposed cultures to either a 1- or 2-µg/cm2 dose of chrysotile for 24 hr before being fixed and stained with anti–-H2AX antibodies. The expression of phosphorylated H2AX as a function of DNA damage was then analyzed using flow cytometry. The histograms represented the frequency of cell number versus the intensity of the fluorescence signals of -H2AX antibody staining [FL1-H (green fluorescence signal received by the photomultiplier tube); Figure 6A]. Even though the number of foci/cell cannot be measured directly by flow cytometry, we found that MEF cells incubated with chrysotile showed an increased staining with anti–-H2AX antibodies as detected by immunofluorescence. However, there was no dose-dependent induction of -H2AX in MEF cells exposed to either 1 or 2 µg/cm2 doses of chrysotile (Figure 6B). Concurrent treatment of catalase greatly suppressed the induction of -H2AX among treated cells. These results suggest that chrysotile induced DNA damage that triggers a stress response leading to H2AX phosphorylation.
Asbestos fiber is an important environmental carcinogen worldwide and remains the primary occupational concern in many developing countries. Although the carcinogenicity of asbestos is well established, the underlying mechanism is not known. We have previously demonstrated that asbestos fibers are mutagenic and induce gene/chromosomal mutations in mammalian cells. Similar results have subsequently been reported by others using various in vitro and in vivo assays that can quantify multilocus deletions (Hei et al. 1992; Lezon-Geyda et al. 1996; Park and Aust 1998). However, it has not been established how asbestos fibers induce such mutational events in vivo.
Inhalation studies in Big Blue lacI transgenic mice have revealed that there is a 1.96-fold increase in mutation frequencies in lung tissues of crocidolite-exposed mice compared with nonexposed control mice, but no specific mutant spectrum has been identified (Rihn et al. 2000). More recently, mutation induction factors ranging from 1.1 to 3.2 in the omenta have been reported in Big Blue lacI transgenic rats injected with crocidolite (Unfried et al. 2002). Intratracheal instillation with amosite results in a 2-fold increase in the mutation frequency in lung DNA in Big Blue lacI transgenic rats (Topinka et al. 2004). It should be noted that the lacI transgenic system is limited to small sequence alterations between 1 and 20 bp, such as point mutations, small deletions, and insertions. Most genome mutations such as large deletions and insertions, translocations, and aneuploidy cannot be effectively recovered by the lacI shuttle vector. Several studies in which mutation frequencies in the lacI transgenic system were compared with that in endogenous genes have shown that spontaneous mutation frequencies at reporter genes were dramatically higher than those found at the endogenous hprt gene (Skopek et al. 1995; Walker et al. 1999). It is likely that overall mutagenesis induced by asbestos fibers may be underestimated in Big Blue lacI mice. As such, it is extremely desirable to establish an efficient system to recover large deletion events induced by asbestos fibers in vivo.
The gpt delta transgenic mouse system, established in the laboratory of one of the co-authors provides a unique opportunity to assess the in vivo mutagenic potential of mineral fibers (Masumura et al. 2003; Nohmi and Masumura 2004, 2005). The gpt delta mice carry tandem repeats of EG10 DNA in two units of 40 phage copies each on both arms of chromosome 17, which are retrievable as phage particles by an in vitro packaging reaction. The rescued phages are then used to quantify the mutation yield upon exposure to genotoxic agents. In the present study the MEF cells from the transgenic mice were used to both quantify and characterize the deletions induced by graded doses of chrysotile fibers. Our results demonstrated that chrysotiles induced a dose dependent increase in mutant yield at the gam and redBA loci in MEF cells and that the incidence and types of mutants generated were comparable to those induced by equitoxic doses of H2O2.
Among the mutants with deletions ? 2 kb that span the redBA/gam gene, the mutant fraction induced by treatment with a 1 µg/cm2 dose of chrysotiles was 5.2-fold higher than those derived spontaneously. The mutant fraction and the number of mutants with deletions > 2 kb, however, were not elevated by further increase in fiber doses. Although the precise reason for this lack of dose–response relationship is not clear, it is possible that mutated cells were selectively killed or that the phages were not effectively recovered in vitro at high fiber doses. In addition to large deletions, the small mutational events observed were predominantly single base pair deletions at the gam locus in both spontaneous mutants and mutants induced by asbestos (Table 1). There is evidence that deletions in the gam gene not only inactivate the gam gene but also interfere with the translation of the redBA gene, leading to functionally inactivate gam and redBA genes(Masumura et al. 2003). It should be noted that the maximum size of deletions detectable by the Spi– assay is 9.6 kb. However, deletions extending into regions adjacent to the transgene concatemer are not detected, as two intact cos sites are required for the packaging of a single vector. Our present study indicated that the maximum deletion generated by chrysotiles in the gpt delta transgenic mutation system were kilobase-sized intrachromosomal deletions, which were much smaller than our previous reports on megabase–sized multilocus deletions generated by asbestos in the human–hamster cells (Hei et al. 1992), largely because of the nature of the model system.
Various in vitro and in vivo studies have indicated that oxyradicals are one of the key determinants of asbestos-induced mutagenesis and carcinogenesis (Shukla et al. 2003). Among the most biologically active oxyradicals (e.g., superoxide anions (O2·), hydroxyl radical (·OH), singlet oxygen (1O2), and hydroperoxy radical (HO2·), H2O2 is relatively long-lived and directly crosses cell membranes by simple diffusion (Root et al. 1975). There is evidence that H2O2 not only induces damage to DNA, causing single- and double-strand breaks, base loss, base substitution, and cross-linking, but also causes chromosome and chromatid aberrations (Mondello et al. 2002). Recently, 8-hydroxydeoxyguanosine, an oxidative DNA damage marker, has been detected in Big Blue lacI transgenic rats treated with asbestos (Unfried et al. 2002). In an effort to understand the molecular mechanisms involved in the intra-chromosomal deletions induced by chrysotile in the present model, we compared mutation patterns between chrysotile asbestos and ROS. In the absence of serum, H2O2 produced predominantly ·OH radicals in human fibroblast culture (Weitzman and Graceffa 1984). Our results showed that for chrysotile-induced mutants the ratios of mutants with large deletions were similar to those induced by H2O2 at equitoxic doses. From a mechanistic point of view, these data suggest that similar mutagenic mechanisms are involved between asbestos fibers and chemically generated ROS. Consistent with this possibility, large mutational events mediated by oxyradicals have been observed in the human–hamster, AS52, and L5178 systems (Fach et al. 2003; Lipinski et al. 2000; Xu et al. 2002).
DSBs are usually regarded as the most deleterious type of DNA damage, induced either by environmental stress, such as irradiation or oxidative stress by the stalling of DNA replication forks (O'Driscoll and Jeggo 2006). Inefficient or inaccurate repair can elevate the frequencies of deletion, amplification, and chromosomal translocation, leading to chromosomal instability and neoplastic transformation. There is evidence that survival fraction and DSB-repair efficiency are dramatically decreased by chrysotile asbestos in the DNA DSB repair deficient cells as compared with wild-type cells (Okayasu et al. 1999). A very early step in the response of mammalian cells to DNA DSBs is the phosphorylation of histone H2AX at serine 139 at the sites of DNA damage (Lowndes and Toh 2005). Using -H2AX as a biomarker for DNA DSBs, our data showed that the accumulation of -H2AX was greatly increased by chrysotile treatment in MEF cells, which was inhibited by concurrent treatment with catalase. These findings provided strong corroborating evidence of the DNA damaging effects of chrysotiles through the oxyradical pathway.
Chromosomal rearrangements have been closely associated with the progression and maintenance of cancer (Radford 2004). One of the major difficulties in detecting in vivo somatic mutations in chromosomal DNA is the lack of systems capable of identifying and isolating mutated genes with high efficiency. Spi– selection based on deletions extending into or through both the redBA and gam genes is an efficient mutation assay system for detecting small to kilobase-sized deletions in different cells, organs, and tissues (Nohmi and Masumura 2004). Although during packaging, the individual genes and vectors are segregated from each other and assayed for mutation independently, the target genes in the gpt delta system are present in multiple copies in tandem arrays and amount to a potential target of approximately 3.8 Mb. In reality megabase deletions cannot be distinguished from kilobase deletions because of the size limitation of lambda phage to be packaged. Thus, it is likely that the deletions that are induced by asbestos fibers in the present study may include intergenic deletions whose sizes are > 10 kb. As gene mutation, mitotic recombination, chromosome loss, and interstitial deletion largely contribute to the development of malignancy, the establishment of the gpt delta transgenic mouse mutation model may provide novel, mechanistic information on asbestos-induced genotoxicity in the future.
|
|
|
| [References Listed in PubMed]
References Bernstein DM, Rogers R, Smith P. 2003. The biopersistence of Canadian chrysotile asbestos following inhalation. Inhal Toxicol 15(13):1247–1274.
Coin PG, Roggli VL, Brody AR. 1994. Persistence of long, thin chrysotile asbestos fibers in the lungs of rats. Environ Health Perspect 102(suppl 5): 197–199.
Dean SW, Brooks TM, Burlinson B, Mirsalis J, Myhr B, Recio L et al. 1999. Transgenic mouse mutation assay systems can play an important role in regulatory mutagenicity testing in vivo for the detection of site-of-contact mutagens. Mutagenesis 14(1):141–151.
Fach E, Kristovich R, Long JF, Waldman WJ, Dutta PK, Williams MV. 2003. The effect of iron on the biological activities of erionite and mordenite. Environ Int 29(4):451–458.
Gardner MJ, Saracci R. 1989. Effects on health of non-occupational exposure to airborne mineral fibres. IARC Sci Publ 90:375–397.
Gustavsson P, Jakobsson R, Johansson H, Lewin F, Norell S, Rutkvist LE. 1998. Occupational exposures and squamous cell carcinoma of the oral cavity, pharynx, larynx, and oesophagus: a case-control study in Sweden. Occup Environ Med 55(6):393–400.
Hei TK, Piao CQ, He ZY, Vannais D, Waldren CA. 1992. Chrysotile fiber is a strong mutagen in mammalian cells. Cancer Res 52(22):6305–6309.
Hogan B, Beddington R, Constantini F, and Lacy E. 1994. Manipulating the Mouse Embryo, 2nd ed. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Horiguchi M, Masumura KI, Ikehata H, Ono T, Kanke Y, Nohmi T. 2001. Molecular nature of ultraviolet B light-induced deletions in the murine epidermis. Cancer Res 61(10):3913–3918.
International Agency for Research on Cancer. 1987. Overall Evaluation of Carcinogenicity: an Updating of IARC Monographs Volumes 1–42. IARC Monogr Eval Carcinog Risks Hum Supplement 7.
Jaurand MC. 1996. Use of in vitro genotoxicity and cell transformation assays to evaluate the potential carcinogenicity of fibres. IARC Sci Publ 140: 55–72.
Khanna KK, Jackson SP. 2001. DNA double-strand breaks: signaling, repair and the cancer connection. Nat Genet 27(3): 247–254.
Kjaerheim K, Ulvestad B, Martinsen JI, Andersen A. 2005. Cancer of the gastrointestinal tract and exposure to asbestos in drinking water among lighthouse keepers (Norway). Cancer Causes Control 16(5):593–598.
Kurose A, Tanaka T, Huang X, Halicka HD, Traganos F, Dai W et al. 2005. Assessment of ATM phosphorylation on Ser-1981 induced by DNA topoisomerase I and II inhibitors in relation to Ser-139-histone H2AX phosphorylation, cell cycle phase, and apoptosis. Cytometry A 68(1):1–9.
Lezon-Geyda K, Jaime CM, Godbold JH, Savransky EF, Hope A, Kheiri SA, et al. 1996. Chrysotile asbestos fibers mediate homologous recombination in Rat2 lambda fibroblasts: implications for carcinogenesis. Mutat Res 361(2–3):113–120.
Lipinski P, Drapier JC, Oliveira L, Retmanska H, Sochanowicz B, Kruszewski M. 2000. Intracellular iron status as a hallmark of mammalian cell susceptibility to oxidative stress: a study of L5178Y mouse lymphoma cell lines differentially sensitive to H2O2. Blood 95(9):2960–2966.
Lowndes NF, Toh GW. 2005. DNA repair: the importance of phosphorylating histone H2AX. Curr Biol 15(3): R99–R102.
Masumura K, Totsuka Y, Wakabayashi K, Nohmi T. 2003. Potent genotoxicity of aminophenylnorharman, formed from non-mutagenic norharman and aniline, in the liver of gpt delta transgenic mouse. Carcinogenesis 24(12):1985–1993.
Miller A. 2005. Mesothelioma in household members of asbestos-exposed workers: 32 United States cases since 1990. Am J Ind Med 47(5):458–462.
Mondello C, Guasconi V, Giulotto E, Nuzzo F. 2002. Gamma-ray and H2O2 induction of gene amplification in hamster cells deficient in DNA double strand break repair. DNA Repair (Amst) 1(6):483–493.
Nohmi T, Katoh M, Suzuki H, Matsui M, Yamada M, Watanabe M, et al. 1996. A new transgenic mouse mutagenesis test system using Spi– and 6-thioguanine selections. Environ Mol Mutagen 28(4):465–470.
Nohmi T, Masumura K. 2005. Molecular nature of intrachromosomal deletions and base substitutions induced by environmental mutagens. Environ Mol Mutagen 45(2–3):150–161.
Nohmi T, Masumura KI. 2004. Gpt delta transgenic mouse: a novel approach for molecular dissection of deletion mutations in vivo. Adv Biophys 38:97–121.
O'Driscoll M, Jeggo PA. 2006. The role of double-strand break repair—insights from human genetics. Nat Rev Genet 7(1):45–54.
Okayasu R, Takahashi S, Yamada S, Hei TK, Ullrich RL. 1999. Asbestos and DNA double strand breaks. Cancer Res 59(2): 298–300.
Pan XL, Day HW, Wang W, Beckett LA, Schenker MB. 2005. Residential proximity to naturally occurring asbestos and mesothelioma risk in California. Am J Respir Crit Care Med 172:1019–1025.
Park SH, Aust AE. 1998. Participation of iron and nitric oxide in the mutagenicity of asbestos in hgprt–, gpt+ Chinese hamster V79 cells. Cancer Res 58(6): 1144–1148.
Poser I, Rahman Q, Lohani M, Yadav S, Becker HH, Weiss DG et al. 2004. Modulation of genotoxic effects in asbestos-exposed primary human mesothelial cells by radical scavengers, metal chelators and a glutathione precursor. Mutat Res 559(1–2): 19–27.
Radford IR. 2004. Chromosomal rearrangement as the basis for human tumourigenesis. Int J Radiat Biol 80(8):543–557.
Rihn B, Coulais C, Kauffer E, Bottin MC, Martin P, Yvon F, et al. 2000. Inhaled crocidolite mutagenicity in lung DNA. Environ Health Perspect 108:341–346.
Rom WN, Hammar SP, Rusch V, Dodson R, Hoffman S. 2001. Malignant mesothelioma from neighborhood exposure to anthophyllite asbestos. Am J Ind Med 40(2): 211–214.
Root RK, Metcalf J, Oshino N, Chance B. 1975. H2O2 release from human granulocytes during phagocytosis. I. Documentation, quantitation, and some regulating factors. J Clin Invest 55(5): 945–955.
Scudiero DA, Shoemaker RH, Paull KD, Monks A, Tierney S, Nofzinger TH, et al. 1988. Evaluation of soluble tetrazolium/ formazan assay for cell growth and drug sensitivity in cul-ture using human and other tumor cell lines. Cancer Res 48:4827–4833.
Shibata A, Kamada N, Masumura K, Nohmi T, Kobayashi S, Teraoka H, et al. 2005. Parp-1 deficiency causes an increase of deletion mutations and insertions/rearrangements in vivo after treatment with an alkylating agent. Oncogene 24(8): 1328–1337.
Shukla A, Gulumian M, Hei TK, Kamp D, Rahman Q, Mossman BT. 2003. Multiple roles of oxidants in the pathogenesis of asbestos-induced diseases. Free Radic Biol Med 34(9):1117–1129.
Skopek TR, Kort KL, Marino DR. 1995. Relative sensitivity of the endogenous hprt gene and lacI transgene in ENU-treated Big Blue B6C3F1 mice. Environ Mol Mutagen 26(1): 9–15.
Timbrell V. 1970. Characteristics of the International Union Against Cancer Standard Reference Samples of Asbestos. In: Pneumoconiosis, Proceedings of the International Conference, April 1969, Johannesburg, South Africa (Shapiro HA, ed). Cape Town, South Africa:Oxford University Press, 28–36.
Topinka J, Loli P, Georgiadis P, Dusinska M, Hurbankova M, Kovacikova Z, et al. 2004. Mutagenesis by asbestos in the lung of lambda-lacI transgenic rats. Mutat Res 553(1–2):67–78.
Unfried K, Schurkes C, Abel J. 2002. Distinct spectrum of mutations induced by crocidolite asbestos: clue for 8-hydroxydeoxyguanosine-dependent mutagenesis in vivo. Cancer Res 62(1):99–104.
Walker VE, Andrews JL, Upton PB, Skopek TR, deBoer JG, Walker DM, et al. 1999. Detection of cyclophosphamide-induced mutations at the Hprt but not the lacI locus in splenic lymphocytes of exposed mice. Environ Mol Mutagen 34(2–3): 167–181.
Weitzman SA, Graceffa P. 1984. Asbestos catalyzes hydroxyl and superoxide radical generation form H2O2. Arch Biochem Biophys 228:373–376.
Xu A, Zhou H, Yu DZ, Hei TK. 2002. Mechanisms of the genotoxicity of crocidolite asbestos in mammalian cells: implication from mutation patterns induced by reactive oxygen species. Environ Health Perspect 110: 1003–1008.
Last Updated: December 13, 2006
|
|
|
|
| |












