Intracellular
Signaling Section
Michael
A Beaven,
B Pharmacy, PhD,
Principal Investigator
Mast cells are best known for their role in
IgE-dependent (atopic) allergic diseases which include allergic
rhinitis, asthma, atopic dermatitis, and anaphylactic reactions
(1). Mast cells are uniquely capable of performing this role
because they express several hundred thousand high affinity receptors
for IgE (FcεR1) and thus respond to IgE-directed antigens.
However, mast cells also express the pathogen-recognizing Toll-like
receptors (TLRs) which probably account for the ability of mast
cells to mount an effective innate immune response to lethal
bacterial infections. Activated mast cells release an array of
potent inflammatory mediators by several mechanisms that include
degranulation with release of preformed mediators, the generation
of inflammatory lipids (eicosanoids) from arachidonic acid, and
robust production of numerous cytokines (Figure 1). Among the
pharmacological options for treatment of mast cell-related inflammatory
diseases, the glucocorticoids remain the most effective drugs
available but they have undesirable side effects (1).
Our objective is to elucidate the biochemical signaling pathways
that mediate the various responses of mast cells to antigen or
TLR ligands. A complementary objective is to understand
how therapeutic agents, especially the glucocorticoids, interrupt
signaling processes so as to provide a rational basis for design
of more selective agents. In fact, the mast cell is a pre-eminent
model for such studies. Most, if not all, known signaling processes
can be activated through the various receptors that are expressed
on mast cells, some of which act in synergy with FcεRI
such as adenosine receptors and Kit (2-4). This permits studies
of the influence of other physiologic factors on antigen-induced
responses.
Past studies highlighted the role of phosphoinositides (5),
calcium (6), and
protein kinase C (7) in degranulation and that of MAP kinases
in the generation of arachidonic acid (8, 9). More recent studies
have focused on the signaling pathways that lead to downstream
activation of transcription factors and production of cytokines
(4) and on the inhibition of these events by glucocorticoids
through induction of inhibitory regulators (10) (Figure 2). Also,
we have revisited a perplexing but key enzyme, phospholipase
D, with respect to its activation (11) and its role in degranulation
(12). Most recent studies have revealed how TLR-ligands act as
potent adjuvants to IgE-mediated reactions which may account
for the exacerbation of allergic diseases by infection (13).
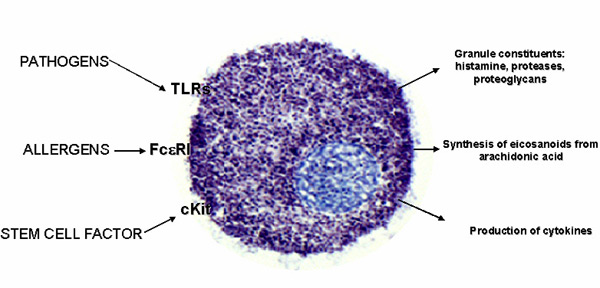
Figure 1. Mast cells are activated primarily through the IgE receptor
(FceRI) by allergens to cause rapid release of granules (stained
blue) that contain histamine, potent proteases, and proteoglycans
such as heparin. Other responses include rapid production of
arachidonic acid-derived prostaglandins and leukotrienes. At
later stages, numerous inflammatory cytokines and chemokines
are produced as a result of gene transcription. As we have shown,
these responses are markedly augmented by pathogenic ligands
of Toll-like receptors (TLR) and the growth factor, Kit ligand
also known as stem cell factor.
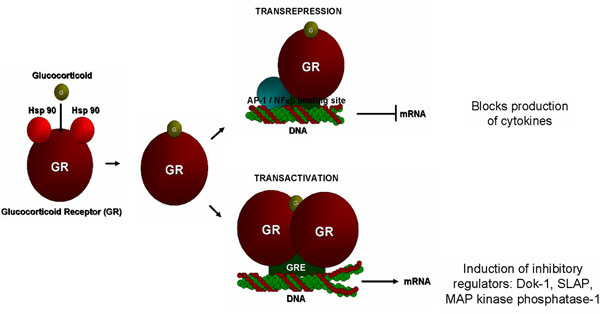 Figure 2. Glucocorticoids act through the
glucocorticoid receptor which may remain as a monomer and thereby
interact with transcription factors to inhibit transcription
of cytokine genes (transrepression) or they can dimerize and
thereby interact with glucocorticoid response elements (GRE)
to induce transcription of genes (transactivation) for metabolic
enzymes and, as we have now shown, inhibitory regulators of signaling
processes in mast cells. The latter observation has broadened
our understanding of the mechanisms of action of glucocorticoids
in allergic disease. Figure 2. Glucocorticoids act through the
glucocorticoid receptor which may remain as a monomer and thereby
interact with transcription factors to inhibit transcription
of cytokine genes (transrepression) or they can dimerize and
thereby interact with glucocorticoid response elements (GRE)
to induce transcription of genes (transactivation) for metabolic
enzymes and, as we have now shown, inhibitory regulators of signaling
processes in mast cells. The latter observation has broadened
our understanding of the mechanisms of action of glucocorticoids
in allergic disease.
References
- Beaven, M. A. and T. R. Hundley.
2003. Mast cell related diseases: Genetics, signaling pathways,
and novel therapies. In ASignal Transduction and Human Disease@
Eds T. Finkel and J.
S. Gutkind. John Wiley & Sons, Hoboken, NJ. pp307-355
- Ali, H., O. H. Choi, K. Yamada, H. M. S. Gonzaga, and M.
A. Beaven. 1996. Sustained activation of phospholipase D via
adenosine A3 receptors is associated with enhancement of antigen-
and Ca2+-ionophore- induced secretion in a rat mast cell line. J.
Pharmacol. Exptl. Therap. 276:837-845
- Beaven, M. A.,
and R. A. Baumgartner. 1996. Downstream
signals initiated in mast cells by FcεRI and other
receptors. Curr.
Opin. Immunol. 8:766-772
- Hundley, T. R., A. M. Gilfillan,
C. Tkaczyk, M. V. Andrade, D. D. Metcalfe, and M. A. Beaven.
2004. Kit and FcεRI
mediate unique and convergent signals for release of inflammatory
mediators from human mast cells. Blood, 104:2410-2417
- Beaven,
M. A., J. P. Moore, G. A. Smith, T. R. Hesketh, and J. C. Metcalfe. 1984. The
calcium signal and phosphatidylinositol breakdown in 2H3 cells. J. Biol. Chem. 259:7137‑7142
- 6)
Beaven, M. A., J. Rogers, J. P. Moore, T. R. Hesketh, G. A.
Smith, and J. C. Metcalfe. 1984. The mechanism
of the calcium signal and correlation with histamine release
in 2H3 cells. J. Biol. Chem. 259:7129‑7136
- Ozawa, K., Z. Szallasi, M. G. Kazanietz, P. M. Blumberg, H.
Mischak, J. F. Mushinski, and M. A. Beaven. 1993. Ca2+‑Dependent
and Ca2+‑independent isozymes of protein kinase C mediate
exocytosis in antigen‑stimulated rat basophilic RBL‑2H3
cells: Reconstitution of secretory responses with Ca2+ and
purified isozymes in washed permeabilized cells. J. Biol.
Chem. 268:1749‑1756
- Hirasawa, N., F.
Santini, and M. A. Beaven. 1995. Activation
of the mitogen‑activated protein kinase/cytosolic phospholipase
A2 pathway in a rat mast cell line by receptor‑dependent
and receptor‑independent stimulants‑Indications
of different pathways for release of arachidonic acid and secretory
granules. J. Immunol. 154:5391-5402.
- Zhang,
C., R. A. Baumgartner, K. Yamada, and M. A. Beaven. 1997. Mitogen
Activated Protein (MAP) Kinase Regulates Production of TNFa
and Release of
Arachidonic Acid in Mast Cells: Indications of Communication
between p38 and p42
MAP kinases. J. Biol. Chem. 272: 13397-13402
- Hiragun,
T. Peng, Z., and M. A. Beaven. 2005. Dexamethasone up-regulates
the inhibitory adaptor protein Dok-1 and suppresses downstream
activation of mitogen-activated protein kinase pathway in antigen-stimulated
RBL-2H3 mast cells. Mol. Pharmacol. 67: 598-603
- Choi, W. S.,
T. Hiragun, J. H. Lee, Y. M. Kim, H.-Y. Kim, A. Chahdi, E.
Her, J. W. Han, and M. A. Beaven. 2004. Activation of RBL-2H3
mast cells is dependent on tyrosine phosphorylation of phospholipase
D2 by Fyn and Fgr. Mol. Cell. Biol. 24:
6980-6992
- Choi, W. S., Y. M. Kim, C. Combs, M. A. Frohman,
and M. A. Beaven. 2002. Phospholipase D1 and D2 regulate
different phases of exocytosis in mast cells. J. Immunol.
168:5682-5689
- Qiao, H., Andrade, M. V., Lisboa, F.
A., Morgan, K., and Beaven, M. A. 2006. FceRI and toll-like
receptors mediate synergistic signals to markedly augment
production of inflammatory cytokines in murine cells. Blood
107: 610-618
|


