5. Hematopoietic Stem Cells
With more than 50 years of experience studying blood-forming stem cells called hematopoietic stem cells, scientists have developed sufficient understanding to actually use them as a therapy. Currently, no other type of stem cell, adult, fetal or embryonic, has attained such status. Hematopoietic stem cell transplants are now routinely used to treat patients with cancers and other disorders of the blood and immune systems. Recently, researchers have observed in animal studies that hematopoietic stem cells appear to be able to form other kinds of cells, such as muscle, blood vessels, and bone. If this can be applied to human cells, it may eventually be possible to use hematopoietic stem cells to replace a wider array of cells and tissues than once thought.
Despite the vast experience with hematopoietic stem cells, scientists face major roadblocks in expanding their use beyond the replacement of blood and immune cells. First, hematopoietic stem cells are unable to proliferate (replicate themselves) and differentiate (become specialized to other cell types) in vitro (in the test tube or culture dish). Second, scientists do not yet have an accurate method to distinguish stem cells from other cells recovered from the blood or bone marrow. Until scientists overcome these technical barriers, they believe it is unlikely that hematopoietic stem cells will be applied as cell replacement therapy in diseases such as diabetes, Parkinson's Disease, spinal cord injury, and many others.
Introduction
Blood cells are responsible for constant maintenance and immune protection of every cell type of the body. This relentless and brutal work requires that blood cells, along with skin cells, have the greatest powers of self-renewal of any adult tissue.
The stem cells that form blood and immune cells are known as hematopoietic stem cells (HSCs). They are ultimately responsible for the constant renewal of blood—the production of billions of new blood cells each day. Physicians and basic researchers have known and capitalized on this fact for more than 50 years in treating many diseases. The first evidence and definition of blood-forming stem cells came from studies of people exposed to lethal doses of radiation in 1945.
Basic research soon followed. After duplicating radiation sickness in mice, scientists found they could rescue the mice from death with bone marrow transplants from healthy donor animals. In the early 1960s, Till and McCulloch began analyzing the bone marrow to find out which components were responsible for regenerating blood [56]. They defined what remain the two hallmarks of an HSC: it can renew itself and it can produce cells that give rise to all the different types of blood cells (see Chapter 4. The Adult Stem Cell).
What Is a Hematopoietic Stem Cell?
A hematopoietic stem cell is a cell isolated from the blood or bone marrow that can renew itself, can differentiate to a variety of specialized cells, can mobilize out of the bone marrow into circulating blood, and can undergo programmed cell death, called apoptosis—a process by which cells that are detrimental or unneeded self-destruct.
A major thrust of basic HSC research since the 1960s has been identifying and characterizing these stem cells. Because HSCs look and behave in culture like ordinary white blood cells, this has been a difficult challenge and this makes them difficult to identify by morphology (size and shape). Even today, scientists must rely on cell surface proteins, which serve, only roughly, as markers of white blood cells.
Identifying and characterizing properties of HSCs began with studies in mice, which laid the groundwork for human studies. The challenge is formidable as about 1 in every 10,000 to 15,000 bone marrow cells is thought to be a stem cell. In the blood stream the proportion falls to 1 in 100,000 blood cells. To this end, scientists began to develop tests for proving the self-renewal and the plasticity of HSCs.
The "gold standard" for proving that a cell derived from mouse bone marrow is indeed an HSC is still based on the same proof described above and used in mice many years ago. That is, the cells are injected into a mouse that has received a dose of irradiation sufficient to kill its own blood-producing cells. If the mouse recovers and all types of blood cells reappear (bearing a genetic marker from the donor animal), the transplanted cells are deemed to have included stem cells.
These studies have revealed that there appear to be two kinds of HSCs. If bone marrow cells from the transplanted mouse can, in turn, be transplanted to another lethally irradiated mouse and restore its hematopoietic system over some months, they are considered to be long-term stem cells that are capable of self-renewal. Other cells from bone marrow can immediately regenerate all the different types of blood cells, but under normal circumstances cannot renew themselves over the long term, and these are referred to as short-term progenitor or precursor cells. Progenitor or precursor cells are relatively immature cells that are precursors to a fully differentiated cell of the same tissue type. They are capable of proliferating, but they have a limited capacity to differentiate into more than one cell type as HSCs do. For example, a blood progenitor cell may only be able to make a red blood cell (see Figure 5.1. Hematopoietic and Stromal Stem Cell Differentiation).
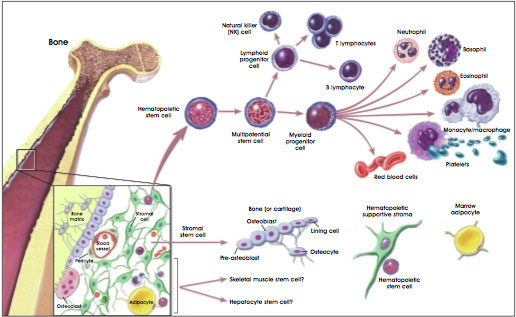
Figure 5.1. Hematopoietic and Stromal Stem Cell Differentiation.
(© 2001 Terese Winslow, Lydia Kibiuk)
Harrison et al. write that short-term blood-progenitor cells in a mouse may restore hematopoiesis for three to four months [36]. The longevity of short-term stem cells for humans is not firmly established. A true stem cell, capable of self-renewal, must be able to renew itself for the entire lifespan of an organism. It is these long-term replicating HSCs that are most important for developing HSC-based cell therapies. Unfortunately, to date, researchers cannot distinguish the long-term from the short-term cells when they are removed from the bloodstream or bone marrow.
The central problem of the assays used to identify long-term stem cells and short-term progenitor cells is that they are difficult, expensive, and time-consuming and cannot be done in humans. A few assays are now available that test cells in culture for their ability to form primitive and long-lasting colonies of cells, but these tests are not accepted as proof that a cell is a long-term stem cell. Some genetically altered mice can receive transplanted human HSCs to test the cells' self-renewal and hematopoietic capabilities during the life of a mouse, but the relevance of this test for the cells in humans—who may live for decades—is open to question.
The difficulty of HSC assays has contributed to two mutually confounding research problems: definitively identifying the HSC and getting it to proliferate, or increase its numbers, in a culture dish. More rapid research progress on characterizing and using HSCs would be possible if they could be readily grown in the laboratory. Conversely, progress in identifying growth conditions suitable for HSCs and getting the cells to multiply would move more quickly if scientists could reliably and readily identify true HSCs.
Can Cell Markers Be Used to Identify Hematopoietic Stem Cells?
HSCs have an identity problem. First, the ones with long-term replicating ability are rare. Second, there are multiple types of stem cells. And, third, the stem cells look like many other blood or bone marrow cells. So how do researchers find the desired cell populations? The most common approach is through markers that appear on the surface of cells. (For a more detailed discussion, see Appendix E.i. Markers: How Do Researchers Use Them to Identify Stem Cells?) These are useful, but not perfect tools for the research laboratory.
In 1988, in an effort to develop a reliable means of identifying these cells, Irving Weissman and his collaborators focused attention on a set of protein markers on the surface of mouse blood cells that were associated with increased likelihood that the cell was a long-term HSC [50]. Four years later, the laboratory proposed a comparable set of markers for the human stem cell [3]. Weissman proposes the markers shown in Table 5.1 as the closest markers for mouse and human HSCs [62].
Table 5.1. Proposed cell-surface markers of undifferentiated hematopoietic stem cells.
| Listed here are cell surface markers found on mouse and human hematopoietic stem cells as they exist in their undifferentiated state in vivo and in vitro. As these cells begin to develop as distinct cell lineages the cell surface markers are no longer identified. |
| Mouse |
Human |
| CD34low/- |
CD 34+ |
| SCA-1+ |
CD59+* |
| Thy1+/low |
Thy1+ |
| CD38+ |
CD38low/- |
| C-kit+ |
C-kit-/low |
| lin-* |
lin-** |
|
* Only one of a family of CD59 markers has thus far been evaluated.
** Lin- cells lack 13 to 14 different mature blood-lineage markers. |
Such cell markers can be tagged with monoclonal antibodies bearing a fluorescent label and culled out of bone marrow with fluorescence-activated cell sorting (FACS).
The groups of cells thus sorted by surface markers are heterogeneous and include some cells that are true, long-term self-renewing stem cells, some shorter-term progenitors, and some non-stem cells. Weissman's group showed that as few as five genetically tagged cells, injected along with larger doses of stem cells into lethally irradiated mice, could establish themselves and produce marked donor cells in all blood cell lineages for the lifetime of the mouse. A single tagged cell could produce all lineages for as many as seven weeks, and 30 purified cells were sufficient to rescue mice and fully repopulate the bone marrow without extra doses of backup cells to rescue the mice [49]. Despite these efforts, researchers remain divided on the most consistently expressed set of HSC markers [27, 32]. Connie Eaves of the University of British Columbia says none of the markers are tied to unique stem cell functions or truly define the stem cell [14]. "Almost every marker I am aware of has been shown to be fickle," she says.
More recently, Diane Krause and her colleagues at Yale University, New York University, and Johns Hopkins University, used a new technique to home in on a single cell capable of reconstituting all blood cell lineages of an irradiated mouse [27]. After marking bone marrow cells from donor male mice with a nontoxic dye, they injected the cells into female recipient mice that had been given a lethal dose of radiation. Over the next two days, some of the injected cells migrated, or homed, to the bone marrow of the recipients and did not divide; when transplanted into a second set of irradiated female mice, they eventually proved to be a concentrated pool of self-renewing stem cells. The cells also reconstituted blood production. The scientists estimate that their technique concentrated the long-term stem cells 500 to 1,000- fold compared with bone marrow.
What Are the Sources of Hematopoietic Stem Cells?
Bone Marrow
The classic source of hematopoietic stem cells (HSCs) is bone marrow. For more than 40 years, doctors performed bone marrow transplants by anesthetizing the stem cell donor, puncturing a bone—typically a hipbone—and drawing out the bone marrow cells with a syringe. About 1 in every 100,000 cells in the marrow is a long-term, blood-forming stem cell; other cells present include stromal cells, stromal stem cells, blood progenitor cells, and mature and maturing white and red blood cells.
Peripheral Blood
As a source of HSCs for medical treatments, bone marrow retrieval directly from bone is quickly fading into history. For clinical transplantation of human HSCs, doctors now prefer to harvest donor cells from peripheral, circulating blood. It has been known for decades that a small number of stem and progenitor cells circulate in the bloodstream, but in the past 10 years, researchers have found that they can coax the cells to migrate from marrow to blood in greater numbers by injecting the donor with a cytokine, such as granulocyte-colony stimulating factor (GCSF). The donor is injected with GCSF a few days before the cell harvest. To collect the cells, doctors insert an intravenous tube into the donor's vein and pass his blood through a filtering system that pulls out CD34+ white blood cells and returns the red blood cells to the donor. Of the cells collected, just 5 to 20 percent will be true HSCs. Thus, when medical researchers commonly refer to peripherally harvested "stem cells," this is something of a misnomer. As is true for bone marrow, the CD34+ cells are a mixture of stem cells, progenitors, and white blood cells of various degrees of maturity.
In the past three years, the majority of autologous (where the donor and recipient are the same person) and allogeneic (where the donor and recipient are different individuals) "bone marrow" transplants have actually been white blood cells drawn from peripheral circulation, not bone marrow. Richard Childs, an intramural investigator at the NIH, says peripheral harvest of cells is easier on the donor—with minimal pain, no anesthesia, and no hospital stay—but also yields better cells for transplants [6]. Childs points to evidence that patients receiving peripherally harvested cells have higher survival rates than bone marrow recipients do. The peripherally harvested cells contain twice as many HSCs as stem cells taken from bone marrow and engraft more quickly. This means patients may recover white blood cells, platelets, and their immune and clotting protection several days faster than they would with a bone marrow graft. Scientists at Stanford report that highly purified, mobilized peripheral cells that have CD34+ and Thy-1+ surface markers engraft swiftly and without complication in breast cancer patients receiving an autologous transplant of the cells after intensive chemotherapy [41].
Umbilical Cord Blood
In the late 1980s and early 1990s, physicians began to recognize that blood from the human umbilical cord and placenta was a rich source of HSCs. This tissue supports the developing fetus during pregnancy, is delivered along with the baby, and, is usually discarded. Since the first successful umbilical cord blood transplants in children with Fanconi anemia, the collection and therapeutic use of these cells has grown quickly. The New York Blood Center's Placental Blood Program, supported by NIH, is the largest U.S. public umbilical cord blood bank and now has 13,000 donations available for transplantation into small patients who need HSCs. Since it began collecting umbilical cord blood in 1992, the center has provided thousands of cord blood units to patients. Umbilical cord blood recipients—typically children—have now lived in excess of eight years, relying on the HSCs from an umbilical cord blood transplant [31, 57].
There is a substantial amount of research being conducted on umbilical cord blood to search for ways to expand the number of HSCs and compare and contrast the biological properties of cord blood with adult bone marrow stem cells. There have been suggestions that umbilical cord blood contains stem cells that have the capability of developing cells of multiple germ layers (multipotent) or even all germ layers, e.g., endoderm, ectoderm, and mesoderm (pluripotent). To date, there is no published scientific evidence to support this claim. While umbilical cord blood represents a valuable resource for HSCs, research data have not conclusively shown qualitative differences in the differentiated cells produced between this source of HSCs and peripheral blood and bone marrow.
Fetal Hematopoietic System
An important source of HSCs in research, but not in clinical use, is the developing blood-producing tissues of fetal animals. Hematopoietic cells appear early in the development of all vertebrates. Most extensively studied in the mouse, HSC production sweeps through the developing embryo and fetus in waves. Beginning at about day 7 in the life of the mouse embryo, the earliest hematopoietic activity is indicated by the appearance of blood islands in the yolk sac (see Appendix A. Early Development). The point is disputed, but some scientists contend that yolk sac blood production is transient and will generate some blood cells for the embryo, but probably not the bulk of the HSCs for the adult animal [12, 26, 44]. According to this proposed scenario, most stem cells that will be found in the adult bone marrow and circulation are derived from cells that appear slightly later and in a different location. This other wave of hematopoietic stem cell production occurs in the AGM—the region where the aorta, gonads, and fetal kidney (mesonephros) begin to develop. The cells that give rise to the HSCs in the AGM may also give rise to endothelial cells that line blood vessels. [13]. These HSCs arise at around days 10 to 11 in the mouse embryo (weeks 4 to 6 in human gestation), divide, and within a couple of days, migrate to the liver [11]. The HSCs in the liver continue to divide and migrate, spreading to the spleen, thymus, and—near the time of birth—to the bone marrow.
Whereas an increasing body of fetal HSC research is emerging from mice and other animals, there is much less information about human fetal and embryonic HSCs. Scientists in Europe, including Coulombel, Peault, and colleagues, first described hematopoietic precursors in human embryos only a few years ago [20, 53]. Most recently, Gallacher and others reported finding HSCs circulating in the blood of 12- to 18-week aborted human fetuses [16, 28, 54] that was rich in HSCs. These circulating cells had different markers than did cells from fetal liver, fetal bone marrow, or umbilical cord blood.
Embryonic Stem Cells and Embryonic Germ Cells
In 1985, it was shown that it is possible to obtain precursors to many different blood cells from mouse embryonic stem cells [9]. Perkins was able to obtain all the major lineages of progenitor cells from mouse embryoid bodies, even without adding hematopoietic growth factors [45].
Mouse embryonic stem cells in culture, given the right growth factors, can generate most, if not all, the different blood cell types [19], but no one has yet achieved the "gold standard" of proof that they can produce long-term HSCs from these sources—namely by obtaining cells that can be transplanted into lethally irradiated mice to reconstitute long-term hematopoiesis [32].
The picture for human embryonic stem and germ cells is even less clear. Scientists from James Thomson's laboratory reported in 1999 that they were able to direct human embryonic stem cells—which can now be cultured in the lab—to produce blood progenitor cells [23]. Israeli scientists reported that they had induced human ES cells to produce hematopoietic cells, as evidenced by their production of a blood protein, gamma-globin [21]. Cell lines derived from human embryonic germ cells (cultured cells derived originally from cells in the embryo that would ultimately give rise to eggs or sperm) that are cultured under certain conditions will produce CD34+ cells [47]. The blood-producing cells derived from human ES and embryonic germ (EG) cells have not been rigorously tested for long-term self-renewal or the ability to give rise to all the different blood cells.
As sketchy as data may be on the hematopoietic powers of human ES and EG cells, blood experts are intrigued by their clinical potential and their potential to answer basic questions on renewal and differentiation of HSCs [19]. Connie Eaves, who has made comparisons of HSCs from fetal liver, cord blood, and adult bone marrow, expects cells derived from embryonic tissues to have some interesting traits. She says actively dividing blood-producing cells from ES cell culture—if they are like other dividing cells—will not themselves engraft or rescue hematopoiesis in an animal whose bone marrow has been destroyed. However, they may play a critical role in developing an abundant supply of HSCs grown in the lab. Indications are that the dividing cells will also more readily lend themselves to gene manipulations than do adult HSCs. Eaves anticipates that HSCs derived from early embryo sources will be developmentally more "plastic" than later HSCs, and more capable of self-renewal [14].
How Do HSCs from Varying Sources Differ?
Scientists in the laboratory and clinic are beginning to measure the differences among HSCs from different sources. In general, they find that HSCs taken from tissues at earlier developmental stages have a greater ability to self-replicate, show different homing and surface characteristics, and are less likely to be rejected by the immune system—making them potentially more useful for therapeutic transplantation.
Stem cell populations of the bone marrow
When do HSCs move from the early locations in the developing fetus to their adult "home" in the bone marrow? European scientists have found that the relative number of CD34+ cells in the collections of cord blood declined with gestational age, but expression of cell-adhesion molecules on these cells increased.
The authors believe these changes reflect preparations for the cells to relocate—from homing in fetal liver to homing in bone marrow [52].
The point is controversial, but a paper by Chen et al. provides evidence that at least in some strains of mice, HSCs from old mice are less able to repopulate bone marrow after transplantation than are cells from young adult mice [5]. Cells from fetal mice were 50 to 100 percent better at repopulating marrow than were cells from young adult mice were. The specific potential for repopulating marrow appears to be strain-specific, but the scientists found this potential declined with age for both strains. Other scientists find no decreases or sometimes increases in numbers of HSCs with age [51]. Because of the difficulty in identifying a long-term stem cell, it remains difficult to quantify changes in numbers of HSCs as a person ages.
Effectiveness of Transplants of Adult versus Umbilical Cord Blood Stem Cells
A practical and important difference between HSCs collected from adult human donors and from umbilical cord blood is simply quantitative. Doctors are rarely able to extract more than a few million HSCs from a placenta and umbilical cord—too few to use in a transplant for an adult, who would ideally get 7 to 10 million CD34+ cells per kilogram body weight, but often adequate for a transplant for a child [33, 48].
Leonard Zon says that HSCs from cord blood are less likely to cause a transplantation complication called graft-versus-host disease, in which white blood cells from a donor attack tissues of the recipient [65]. In a recent review of umbilical cord blood transplantation, Laughlin cites evidence that cord blood causes less graft-versus-host disease [31]. Laughlin writes that it is yet to be determined whether umbilical cord blood HSCs are, in fact, longer lived in a transplant recipient.
In lab and mouse-model tests comparing CD34+ cells from human cord with CD34+ cells derived from adult bone marrow, researchers found cord blood had greater proliferation capacity [24]. White blood cells from cord blood engrafted better in a mouse model, which was genetically altered to tolerate the human cells, than did their adult counterparts.
Effectiveness in Transplants of Peripheral Versus Bone Marrow Stem Cells
In addition to being far easier to collect, peripherally harvested white blood cells have other advantages over bone marrow. Cutler and Antin's review says that peripherally harvested cells engraft more quickly, but are more likely to cause graft-versus-host disease [8]. Prospecting for the most receptive HSCs for gene therapy, Orlic and colleagues found that mouse HSCs mobilized with cytokines were more likely to take up genes from a viral vector than were non-mobilized bone marrow HSCs [43].
What Do Hematopoietic Stem Cells Do and What Factors Are Involved in These Activities?
As stated earlier, an HSC in the bone marrow has four actions in its repertoire: 1) it can renew itself, 2) it can differentiate, 3) it can mobilize out of the bone marrow into circulation (or the reverse), or 4) it can undergo programmed cell death, or apoptosis. Understanding the how, when, where, which, and why of this simple repertoire will allow researchers to manipulate and use HSCs for tissue and organ repair.
Self-renewal of Hematopoietic Stem Cells
Scientists have had a tough time trying to grow—or even maintain—true stem cells in culture. This is an important goal because cultures of HSCs that could maintain their characteristic properties of self-renewal and lack of differentiation could provide an unlimited source of cells for therapeutic transplantation and study. When bone marrow or blood cells are observed in culture, one often observes large increases in the number of cells. This usually reflects an increase in differentiation of cells to progenitor cells that can give rise to different lineages of blood cells but cannot renew themselves. True stem cells divide and replace themselves slowly in adult bone marrow.
New tools for gene-expression analysis will now allow scientists to study developmental changes in telomerase activity and telomeres. Telomeres are regions of DNA found at the end of chromosomes that are extended by the enzyme telomerase. Telomerase activity is necessary for cells to proliferate and activity decreases with age leading to shortened telomeres. Scientists hypothesize that declines in stem cell renewal will be associated with declines in telomere length and telomerase activity. Telomerase activity in hematopoietic cells is associated with self-renewal potential [40].
Because self-renewal divisions are rare, hard to induce in culture, and difficult to prove, scientists do not have a definitive answer to the burning question: what puts—or perhaps keeps—HSCs in a self-renewal division mode? HSCs injected into an anemic patient or mouse—or one whose HSCs have otherwise been suppressed or killed—will home to the bone marrow and undergo active division to both replenish all the different types of blood cells and yield additional self-renewing HSCs. But exactly how this happens remains a mystery that scientists are struggling to solve by manipulating cultures of HSCs in the laboratory.
Two recent examples of progress in the culturing studies of mouse HSCs are by Ema and coworkers and Audet and colleagues [2, 15]. Ema et al. found that two cytokines—stem cell factor and thrombo-poietin—efficiently induced an unequal first cell division in which one daughter cell gave rise to repopulating cells with self-renewal potential. Audet et al. found that activation of the signaling molecule gp130 is critical to survival and proliferation of mouse HSCs in culture.
Work with specific cytokines and signaling molecules builds on several earlier studies demonstrating modest increases in the numbers of stem cells that could be induced briefly in culture. For example, Van Zant and colleagues used continuous-perfusion culture and bioreactors in an attempt to boost human HSC numbers in single cord blood samples incubated for one to two weeks [58]. They obtained a 20-fold increase in "long-term culture initiating cells."
More clues on how to increase numbers of stem cells may come from looking at other animals and various developmental stages. During early developmental stages—in the fetal liver, for example—HSCs may undergo more active cell division to increase their numbers, but later in life, they divide far less often [30, 42]. Culturing HSCs from 10- and 11-day-old mouse embryos, Elaine Dzierzak at Erasmus University in the Netherlands finds she can get a 15-fold increase in HSCs within the first 2 or 3 days after she removes the AGM from the embryos [38]. Dzierzak recognizes that this is dramatically different from anything seen with adult stem cells and suggests it is a difference with practical importance. She suspects that the increase is not so much a response to what is going on in the culture but rather, it represents the developmental momentum of this specific embryonic tissue. That is, it is the inevitable consequence of divisions that were cued by that specific embryonic microenvironment. After five days, the number of HSCs plateaus and can be maintained for up to a month. Dzierzak says that the key to understanding how adult-derived HSCs can be expanded and manipulated for clinical purposes may very well be found by defining the cellular composition and complex molecular signals in the AGM region during development [13].
In another approach, Lemischka and coworkers have been able to maintain mouse HSCs for four to seven weeks when they are grown on a clonal line of cells (AFT024) derived from the stroma, the other major cellular constituent of bone marrow [39]. No one knows which specific factors secreted by the stromal cells maintain the stem cells. He says ongoing gene cloning is rapidly zeroing in on novel molecules from the stromal cells that may "talk" to the stem cells and persuade them to remain stem cells—that is, continue to divide and not differentiate.
If stromal factors provide the key to stem cell self-renewal, research on maintaining stromal cells may be an important prerequisite. In 1999, researchers at Osiris Therapeutics and Johns Hopkins University reported culturing and expanding the numbers of mesenchymal stem cells, which produce the stromal environment [46]. Whereas cultured HSCs rush to differentiate and fail to retain primitive, self-renewing cells, the mesenchymal stem cells could be increased in numbers and still retained their powers to generate the full repertoire of descendant lineages.
Differentiation of HSCs into Components of the Blood and Immune System
Producing differentiated white and red blood cells is the real work of HSCs and progenitor cells. M.C. MacKey calculates that in the course of producing a mature, circulating blood cell, the original hematopoietic stem cell will undergo between 17 and 19.5 divisions, "giving a net amplification of between ~170,000 and ~720,000" [35].
Through a series of careful studies of cultured cells—often cells with mutations found in leukemia patients or cells that have been genetically altered—investigators have discovered many key growth factors and cytokines that induce progenitor cells to make different types of blood cells. These factors interact with one another in complex ways to create a system of exquisite genetic control and coordination of blood cell production.
Migration of Hematopoietic Stem Cells Into and Out of Marrow and Tissues
Scientists know that much of the time, HSCs live in intimate connection with the stroma of bone marrow in adults (see Chapter 4. The Adult Stem Cell). But HSCs may also be found in the spleen, in peripheral blood circulation, and other tissues. Connection to the interstices of bone marrow is important to both the engraftment of transplanted cells and to the maintenance of stem cells as a self-renewing population. Connection to stroma is also important to the orderly proliferation, differentiation, and maturation of blood cells [63].
Weissman says HSCs appear to make brief forays out of the marrow into tissues, then duck back into marrow [62]. At this time, scientists do not understand why or how HSCs leave bone marrow or return to it [59]. Scientists find that HSCs that have been mobilized into peripheral circulation are mostly non-dividing cells [64]. They report that adhesion molecules on the stroma, play a role in mobilization, in attachment to the stroma, and in transmitting signals that regulate HSC self-renewal and progenitor differentiation [61].
Apoptosis and Regulation of Hematopoietic Stem Cell Populations
The number of blood cells in the bone marrow and blood is regulated by genetic and molecular mechanisms. How do hematopoietic stem cells know when to stop proliferating? Apoptosis is the process of programmed cell death that leads cells to self-destruct when they are unneeded or detrimental. If there are too few HSCs in the body, more cells divide and boost the numbers. If excess stem cells were injected into an animal, they simply wouldn't divide or would undergo apoptosis and be eliminated [62]. Excess numbers of stem cells in an HSC transplant actually seem to improve the likelihood and speed of engraftment, though there seems to be no rigorous identification of a mechanism for this empirical observation.
The particular signals that trigger apoptosis in HSCs are as yet unknown. One possible signal for apoptosis might be the absence of life-sustaining signals from bone marrow stroma. Michael Wang and others found that when they used antibodies to disrupt the adhesion of HSCs to the stroma via VLA-4/VCAM-1, the cells were predisposed to apoptosis [61].
Understanding the forces at play in HSC apoptosis is important to maintaining or increasing their numbers in culture. For example, without growth factors, supplied in the medium or through serum or other feeder layers of cells, HSCs undergo apoptosis. Domen and Weissman found that stem cells need to get two growth factor signals to continue life and avoid apoptosis: one via a protein called BCL-2, the other from steel factor, which, by itself, induces HSCs to produce progenitor cells but not to self-renew [10].
What Are the Clinical Uses of Hematopoietic Stem Cells?
Leukemia and Lymphoma
Among the first clinical uses of HSCs were the treatment of cancers of the blood—leukemia and lymphoma, which result from the uncontrolled proliferation of white blood cells. In these applications, the patient's own cancerous hematopoietic cells were destroyed via radiation or chemotherapy, then replaced with a bone marrow transplant, or, as is done now, with a transplant of HSCs collected from the peripheral circulation of a matched donor. A matched donor is typically a sister or brother of the patient who has inherited similar human leukocyte antigens (HLAs) on the surface of their cells. Cancers of the blood include acute lymphoblastic leukemia, acute myeloblastic leukemia, chronic myelogenous leukemia (CML), Hodgkin's disease, multiple myeloma, and non-Hodgkin's lymphoma.
Thomas and Clift describe the history of treatment for chronic myeloid leukemia as it moved from largely ineffective chemotherapy to modestly successful use of a cytokine, interferon, to bone marrow trans-plants—first in identical twins, then in HLA-matched siblings [55]. Although there was significant risk of patient death soon after the transplant either from infection or from graft-versus-host disease, for the first time, many patients survived this immediate challenge and had survival times measured in years or even decades, rather than months. The authors write, "In the space of 20 years, marrow transplantation has contributed to the transformation of [chronic myelogenous leukemia] CML from a fatal disease to one that is frequently curable. At the same time, experience acquired in this setting has improved our understanding of many transplant-related problems. It is now clear that morbidity and mortality are not inevitable consequences of allogeneic transplantation, [and] that an allogeneic effect can add to the anti-leukemic power of conditioning regimens…"
In a recent development, CML researchers have taken their knowledge of hematopoietic regulation one step farther. On May 10, 2001, the Food and Drug Administration approved Gleevec™ (imatinib mesylate), a new, rationally designed oral drug for treatment of CML. The new drug specifically targets a mutant protein, produced in CML cancer cells, that sabotages the cell signals controlling orderly division of progenitor cells. By silencing this protein, the new drug turns off cancerous overproduction of white blood cells, so doctors do not have to resort to bone marrow transplantation. At this time, it is unknown whether the new drug will provide sustained remission or will prolong life for CML patients.
Inherited Blood Disorders
Another use of allogeneic bone marrow transplants is in the treatment of hereditary blood disorders, such as different types of inherited anemia (failure to produce blood cells), and inborn errors of metabolism (genetic disorders characterized by defects in key enzymes need to produce essential body components or degrade chemical byproducts). The blood disorders include aplastic anemia, beta-thalassemia, Blackfan-Diamond syndrome, globoid cell leukodystrophy, sickle-cell anemia, severe combined immunodeficiency, X-linked lymphoproliferative syndrome, and Wiskott-Aldrich syndrome. Inborn errors of metabolism that are treated with bone marrow transplants include: Hunter's syndrome, Hurler's syndrome, Lesch Nyhan syndrome, and osteopetrosis. Because bone marrow transplantation has carried a significant risk of death, this is usually a treatment of last resort for otherwise fatal diseases.
Hematopoietic Stem Cell Rescue in Cancer Chemotherapy
Chemotherapy aimed at rapidly dividing cancer cells inevitably hits another target—rapidly dividing hematopoietic cells. Doctors may give cancer patients an autologous stem cell transplant to replace the cells destroyed by chemotherapy. They do this by mobilizing HSCs and collecting them from peripheral blood. The cells are stored while the patient undergoes intensive chemotherapy or radiotherapy to destroy the cancer cells. Once the drugs have washed out of a patient's body, the patient receives a transfusion of his or her stored HSCs. Because patients get their own cells back, there is no chance of immune mismatch or graft-versus-host disease. One problem with the use of autologous HSC transplants in cancer therapy has been that cancer cells are sometimes inadvertently collected and reinfused back into the patient along with the stem cells. One team of investigators finds that they can prevent reintroducing cancer cells by purifying the cells and preserving only the cells that are CD34+, Thy-1+[41].
Graft-Versus-Tumor Treatment of Cancer
One of the most exciting new uses of HSC transplantation puts the cells to work attacking otherwise untreatable tumors. A group of researchers in NIH's intramural research program recently described this approach to treating metastatic kidney cancer [7]. Just under half of the 38 patients treated so far have had their tumors reduced. The research protocol is now expanding to treatment of other solid tumors that resist standard therapy, including cancer of the lung, prostate, ovary, colon, esophagus, liver, and pancreas.
This experimental treatment relies on an allogeneic stem cell transplant from an HLA-matched sibling whose HSCs are collected peripherally. The patient's own immune system is suppressed, but not totally destroyed. The donor's cells are transfused into the patient, and for the next three months, doctors closely monitor the patient's immune cells, using DNA fingerprinting to follow the engraftment of the donor's cells and regrowth of the patient's own blood cells. They must also judiciously suppress the patient's immune system as needed to deter his/her T cells from attacking the graft and to reduce graft-versus-host disease.
A study by Joshi et al. shows that umbilical cord blood and peripherally harvested human HSCs show antitumor activity in the test tube against leukemia cells and breast cancer cells [22]. Grafted into a mouse model that tolerates human cells, HSCs attack human leukemia and breast cancer cells. Although untreated cord blood lacks natural killer (NK) lymphocytes capable of killing tumor cells, researchers have found that at least in the test tube and in mice, they can greatly enhance the activity and numbers of these cells with cytokines IL-15 [22, 34].
Other Applications of Hematopoietic Stem Cells
Substantial basic and limited clinical research exploring the experimental uses of HSCs for other diseases is underway. Among the primary applications are autoimmune diseases, such as diabetes, rheumatoid arthritis, and system lupus erythematosis. Here, the body's immune system turns to destroying body tissues. Experimental approaches similar to those applied above for cancer therapies are being conducted to see if the immune system can be reconstituted or reprogrammed. More detailed discussion on this application is provided in Chapter 6. Autoimmune Diseases and the Promise of Stem Cell-Based Therapies. The use of HSCs as a means to deliver genes to repair damaged cells is another application being explored. The use of HSCs for gene therapies is discussed in detail in Chapter 11. Use of Genetically Modified Stem Cells in Experimental Gene Therapies.
Plasticity of Hematopoietic Stem Cells
A few recent reports indicate that scientists have been able to induce bone marrow or HSCs to differentiate into other types of tissue, such as brain, muscle, and liver cells. These concepts and the experimental evidence supporting this concept are discussed in Chapter 4. The Adult Stem Cell.
Research in a mouse model indicates that cells from grafts of bone marrow or selected HSCs may home to damaged skeletal and cardiac muscle or liver and regenerate those tissues [4, 29]. One recent advance has been in the study of muscular dystrophy, a genetic disease that occurs in young people and leads to progressive weakness of the skeletal muscles. Bittner and colleagues used mdx mice, a genetically modified mouse with muscle cell defects similar to those in human muscular dystrophy. Bone marrow from non-mdx male mice was transplanted into female mdx mice with chronic muscle damage; after 70 days, researchers found that nuclei from the males had taken up residence in skeletal and cardiac muscle cells.
Lagasse and colleagues' demonstration of liver repair by purified HSCs is a similarly encouraging sign that HSCs may have the potential to integrate into and grow in some non-blood tissues. These scientists lethally irradiated female mice that had an unusual genetic liver disease that could be halted with a drug. The mice were given transplants of genetically marked, purified HSCs from male mice that did not have the liver disease. The transplants were given a chance to engraft for a couple of months while the mice were on the liver-protective drug. The drug was then removed, launching deterioration of the liver—and a test to see whether cells from the transplant would be recruited and rescue the liver. The scientists found that transplants of as few as 50 cells led to abundant growth of marked, donor-derived liver cells in the female mice.
Recently, Krause has shown in mice that a single selected donor hematopoietic stem cell could do more than just repopulate the marrow and hematopoietic system of the recipient [27]. These investigators also found epithelial cells derived from the donors in the lungs, gut, and skin of the recipient mice. This suggests that HSCs may have grown in the other tissues in response to infection or damage from the irradiation the mice received.
In humans, observations of male liver cells in female patients who have received bone marrow grafts from males, and in male patients who have received liver transplants from female donors, also suggest the possibility that some cells in bone marrow have the capacity to integrate into the liver and form hepatocytes [1].
What Are the Barriers to the Development of New and Improved Treatments Using Hematopoietic Stem Cells?
Boosting the Numbers of Hematopoietic Stem Cells
Clinical investigators share the same fundamental problem as basic investigators—limited ability to grow and expand the numbers of human HSCs. Clinicians repeatedly see that larger numbers of cells in stem cell grafts have a better chance of survival in a patient than do smaller numbers of cells. The limited number of cells available from a placenta and umbilical cord blood transplant currently means that cord blood banks are useful to pediatric but not adult patients. Investigators believe that the main cause of failure of HSCs to engraft is host-versus-graft disease, and larger grafts permit at least some donor cells to escape initial waves of attack from a patient's residual or suppressed immune system [6]. Ability to expand numbers of human HSCs in vivo or in vitro would clearly be an enormous boost to all current and future medical uses of HSC transplantation.
Once stem cells and their progeny can be multiplied in culture, gene therapists and blood experts could combine their talents to grow limitless quantities of "universal donor" stem cells, as well as progenitors and specific types of red and white blood cells. If the cells were engineered to be free of markers that provoke rejection, these could be transfused to any recipient to treat any of the diseases that are now addressed with marrow, peripheral, cord, or other transfused blood. If gene therapy and studies of the plasticity of HSCs succeed, the cells could also be grown to repair other tissues and treat non-blood-related disorders [32].
Several research groups in the United States, Canada, and abroad have been striving to find the key factor or factors for boosting HSC production. Typical approaches include comparing genes expressed in primitive HSCs versus progenitor cells; comparing genes in actively dividing fetal HSCs versus adult HSCs; genetic screening of hematopoietically mutated zebrafish; studying dysregulated genes in cancerous hematopoietic cells; analyzing stromal or feeder-layer factors that appear to boost HSC division; and analyzing factors promoting homing and attachment to the stroma. Promising candidate factors have been tried singly and in combination, and researchers claim they can now increase the number of long-term stem cells 20-fold, albeit briefly, in culture.
The specific assays researchers use to prove that their expanded cells are stem cells vary, which makes it difficult to compare the claims of different research groups. To date, there is only a modest ability to expand true, long-term, self-renewing human HSCs. Numbers of progenitor cells are, however, more readily increased. Kobari et al., for example, can increase progenitor cells for granulocytes and macrophages 278-fold in culture [25].
Some investigators are now evaluating whether these comparatively modest increases in HSCs are clinically useful. At this time, the increases in cell numbers are not sustainable over periods beyond a few months, and the yield is far too low for mass production. In addition, the cells produced are often not rigorously characterized. A host of other questions remain—from how well the multiplied cells can be altered for gene therapy to their potential longevity, immunogenicity, ability to home correctly, and susceptibility to cancerous transformation. Glimm et al. [17] highlight some of these problems, for example, with their confirmation that human stem cells lose their ability to repopulate the bone marrow as they enter and progress through the cell cycleælike mouse stem cells that have been stimulated to divide lose their transplantability [18]. Observations on the inverse relationship between progenitor cell division rate and longevity in strains of mice raise an additional concern that culture tricks or selection of cells that expand rapidly may doom the cells to a short life.
Pragmatically, some scientists say it may not be necessary to be able to induce the true, long-term HSC to divide in the lab. If they can manipulate progenitors and coax them into division on command, gene uptake, and differentiation into key blood cells and other tissues, that may be sufficient to accomplish clinical goals. It might be sufficient to boost HSCs or subpopulations of hematopoietic cells within the body by chemically prodding the bone marrow to supply the as-yet-elusive factors to rejuvenate cell division.
Outfoxing the Immune System in Host, Graft, and Pathogen Attacks
Currently, the risks of bone marrow transplants—graft rejection, host-versus-graft disease, and infection during the period before HSCs have engrafted and resumed full blood cell production—restrict their use to patients with serious or fatal illnesses. Allogeneic grafts must come from donors with a close HLA match to the patient (see Chapter 6. Autoimmune Diseases and the Promise of Stem Cell-Based Therapies). If doctors could precisely manipulate immune reactions and protect patients from pathogens before their transplants begin to function, HSC transplants could be extended to less ill patients and patients for whom the HLA match was not as close as it must now be. Physicians might use transplants with greater impunity in gene therapy, autoimmune disease, HIV/AIDS treatment, and the preconditioning of patients to accept a major organ transplant.
Scientists are zeroing in on subpopulations of T cells that may cause or suppress potentially lethal host-versus-graft rejection and graft-versus-host disease in allogeneic-transplant recipients. T cells in a graft are a two-edged sword. They fight infections and help the graft become established, but they also can cause graft-versus-host disease. Identifying subpopulations of T cells responsible for deleterious and beneficial effects—in the graft, but also in residual cells surviving or returning in the host—could allow clinicians to make grafts safer and to ratchet up graft-versus-tumor effects [48]. Understanding the presentation of antigens to the immune system and the immune system's healthy and unhealthy responses to these antigens and maturation and programmed cell death of T cells is crucial.
The approach taken by investigators at Stanford—purifying peripheral blood—may also help eliminate the cells causing graft-versus-host disease. Transplants in mouse models support the idea that purified HSCs, cleansed of mature lymphocytes, engraft readily and avoid graft-versus-host disease [60].
Knowledge of the key cellular actors in autoimmune disease, immune grafting, and graft rejection could also permit scientists to design gentler "minitransplants." Rather than obliterating and replacing the patient's entire hematopoietic system, they could replace just the faulty components with a selection of cells custom tailored to the patient's needs. Clinicians are currently experimenting with deletion of T cells from transplants in some diseases, for example, thereby reducing graft-versus-host disease.
Researchers are also experimenting with the possibility of knocking down the patient's immune system—but not knocking it out. A blow that is sublethal to the patient's hematopoietic cells given before an allogeneic transplant can be enough to give the graft a chance to take up residence in the bone marrow. The cells replace some or all of the patient's original stem cells, often making their blood a mix of donor and original cells. For some patients, this mix of cells will be enough to accomplish treatment objectives but without subjecting them to the vicious side effects and infection hazards of the most powerful treatments used for total destruction of their hematopoietic systems [37].
Understanding the Differentiating Environment and Developmental Plasticity
At some point in embryonic development, all cells are plastic, or developmentally flexible enough to grow into a variety of different tissues. Exactly what is it about the cell or the embryonic environment that instructs cells to grow into one organ and not another?
Could there be embryological underpinnings to the apparent plasticity of adult cells? Researchers have suggested that a lot of the tissues that are showing plasticity are adjacent to one another after gastrulation in the sheet of mesodermal tissue that will go on to form blood—muscle, blood vessels, kidney, mesenchyme, and notochord. Plasticity may reflect derivation from the mesoderm, rather than being a fixed trait of hematopoietic cells. One lab is now studying the adjacency of embryonic cells and how the developing embryo makes the decision to make one tissue instead of another—and whether the decision is reversible [65].
In vivo studies of the plasticity of bone marrow or purified stem cells injected into mice are in their infancy. Even if follow-up studies confirm and more precisely characterize and quantify plasticity potential of HSCs in mice, there is no guarantee that it will occur or can be induced in humans.
Summary
Grounded in half a century of research, the study of hematopoietic stem cells is one of the most exciting and rapidly advancing disciplines in biomedicine today. Breakthrough discoveries in both the laboratory and clinic have sharply expanded the use and supply of life-saving stem cells. Yet even more promising applications are on the horizon and scientists' current inability to grow HSCs outside the body could delay or thwart progress with these new therapies. New treatments include graft-versus-tumor therapy for currently incurable cancers, autologous transplants for autoimmune diseases, and gene therapy and tissue repair for a host of other problems. The techniques, cells, and knowledge that researchers have now are inadequate to realize the full promise of HSC-based therapy.
Key issues for tapping the potential of hematopoietic stem cells will be finding ways to safely and efficiently expand the numbers of transplantable human HSCs in vitro or in vivo. It will also be important to gain a better understanding of the fundamentals of how immune cells work—in fighting infections, in causing transplant rejection, and in graft-versus-host disease as well as master the basics of HSC differentiation. Concomitant advances in gene therapy techniques and the understanding of cellular plasticity could make HSCs one of the most powerful tools for healing.
References
- Alison, M.R., Poulsom, R., Jeffery, R., Dhillon, A.P., Quaglia, A., Jacob, J., Novelli, M., Prentice, G., Williamson, J., and Wright, N.A. (2000). Hepatocytes from non-hepatic adult stem cells. Nature. 406, 257.
- Audet, J., Miller, C.L., Rose-John, S., Piret, J.M., and Eaves, C.J. (2001). Distinct role of gp130 activation in promotingself-renewal divisions by mitogenically stimulated murine hematopoietic stem cells. Proc. Natl. Acad. Sci. U. S. A. 98, 1757–1762.
- Baum, C.M., Weissman, I.L., Tsukamoto, A.S., Buckle, A.M., and Peault, B. (1992). Isolation of a candidate human hematopoietic stem-cell population. Proc. Natl. Acad. Sci. U. S. A. 89, 2804–2808.
- Bittner, R.E., Schofer, C., Weipoltshammer, K., Ivanova, S., Streubel, B., Hauser, E., Freilinger, M., Hoger, H., Elbe-Burger, A., and Wachtler, F. (1999). Recruitment of bone-marrow-derived cells by skeletal and cardiac muscle in adult dystrophic mdx mice. Anat. Embryol. (Berl) 199, 391–396.
- Chen, J., Astle, C.M., and Harrison, D.E. (1999). Development and aging of primitive hematopoietic stem cells in BALB/cBy mice. Exp. Hematol. 27, 928–935.
- Childs, R., personal communication.
- Childs, R., Chernoff, A., Contentin, N., Bahceci, E., Schrump, D., Leitman, S., Read, E.J., Tisdale, J., Dunbar, C., Linehan, W.M., Young, N.S., and Barrett, A.J. (2000). Regression of metastatic renal-cell carcinoma after nonmyeloablative allogeneic peripheral-blood stem-cell transplantation. N. Engl. J. Med. 343, 750–758.
- Cutler, C. and Antin, J.H. (2001). Peripheral blood stem cells for allogeneic transplantation: a review. Stem Cells. 19, 108–117.
- Doetschman, T., Eistetter, H., Katz, M., Schmit, W., and Kemler, R. (1985). The in vitro development of blastocyst-derived embryonic stem cell lines: formation of visceral yolk sac, blood islands and myocardium. J. Embryol. Exp. Morph. 87, 27–45.
- Domen, J. and Weissman, I.L. (2000). Hematopoietic stem cells need two signals to prevent apoptosis; BCL-2 can provide one of these, Kitl/c-Kit signaling the other. J. Exp. Med. 192, 1707–1718.
- Dzierzak, E., Medvinsky, A., and de Bruijn, M. (1998). Qualitative and quantitative aspects of haematopoietic cell development in the mammalian embryo. Immunol. Today. 19, 228–236.
- Dzierzak, E. (1999). Embryonic beginnings of definitive hematopoietic stem cells. Ann. N. Y. Acad. Sci. 872, 256–262.
- Dzierzak, E., personal communication.
- Eaves, C. J., personal communication.
- Ema, H., Takano, H., Sudo, K., and Nakauchi, H. (2000). In vitro self-renewal division of hematopoietic stem cells. J. Exp. Med. 192, 1281–1288.
- Gallacher, L., Murdoch, B., Wu, D., Karanu, F., Fellows, F., and Bhatia, M. (2000). Identification of novel circulating human embryonic blood stem cells. Blood. 96, 1740–1747.
- Glimm, H., Oh, I.H., and Eaves, C.J. (2000). Human hematopoietic stem cells stimulated to proliferate in vitro lose engraftment potential during their S/G(2)/M transit and do not reenter G(0). Blood. 96, 4185–4193.
- Gothot, A., van der Loo, J.C., Clapp, D.W., and Srour, E.F. (1998). Cell cycle-related changes in repopulating capacity of human mobilized peripheral blood CD34+ cells in non-obese diabetic/severe combined immune-deficient mice. Blood. 92, 2641–2649.
- Hole, N. (1999). Embryonic stem cell-derived haematopoiesis. Cells Tissues Organs. 165, 181–189.
- Huyhn, A., Dommergues, M., Izac, B., Croisille, L., Katz, A., Vainchenker, W., and Coulombel, L. (1995). Characterization of hematopoietic progenitors from human yolk sacs and embryos. Blood. 86, 4474–4485.
- Itskovitz-Eldor, J., Schuldiner, M., Karsenti, D., Eden, A., Yanuka, O., Amit, M., Soreq, H., and Benvenisty, N. (2000). Differentiation of human embryonic stem cells into embryoid bodies comprising the three embryonic germ layers. Mol. Med. 6, 88–95.
- Joshi, S.S., Tarantolo, S.R., Kuszynski, C.A., and Kessinger, A. (2000). Antitumor therapeutic potential of activated human umbilical cord blood cells against leukemia and breast cancer. Clin. Cancer Res. 6, 4351–4358.
- Kaufman, D.S., Lewis, R.L., Auerbach, R., and Thomson, J.A. (1999). Directed differentiation of human embryonic stem cells into hematopoietic colony forming cells. Blood. 94 (Supplement part 1), 34a.
- Kim, D.K., Fujiki, Y., Fukushima, T., Ema, H., Shibuya, A., and Nakauchi, H. (1999). Comparison of hematopoietic activities of human bone marrow and umbilical cord blood CD34 positive and negative cells. Stem Cells. 17, 286–294.
- Kobari, L., Pflumio, F., Giarratana, M., Li, X., Titeux, M., Izac, B., Leteurtre, F., Coulombel, L., and Douay, L. (2000). In vitro and in vivo evidence for the long-term multilineage (myeloid, B, NK, and T) reconstitution capacity of ex vivo expanded human CD34+ cord blood cells. Exp. Hematol. 28, 1470–1480.
- Koichi, T., Akashi, K., and Weissman, I.L. (2001). Stem cells and hematolymphoic development. Zon, L.I. ed. Oxford Press.
- Krause, D.S., Theise, N.D., Collector, M.I., Henegariu, O., Hwang, S., Gardner, R., Neutzel, S., and Sharkis, S.J. (2001). Multi-organ, multi-lineage engraftment by a single bone marrow-derived stem cell. Cell. 105, 369–377.
- Labastie, M.C., Cortes, F., Romeo, P.H., Dulac, C., and Peault, B. (1998). Molecular identity of hematopoietic precursor cells emerging in the human embryo. Blood. 92, 3624–3635.
- Lagasse, E., Connors, H., Al Dhalimy, M., Reitsma, M., Dohse, M., Osborne, L., Wang, X., Finegold, M., Weissman, I.L., and Grompe, M. (2000). Purified hematopoietic stem cells can differentiate into hepatocytes in vivo. Nat. Med. 6, 1229–1234.
- Lansdorp, P.M., Dragowska, W., and Mayani, H. (1993). Ontogeny-related changes in proliferative potential of human hematopoietic cells. J. Exp. Med. 178, 787–791.
- Laughlin, M.J. (2001). Umbilical cord blood for allogeneic transplantation in children and adults. Bone Marrow Transplant. 27, 1–6.
- Lemischka, I. R., personal communication.
- Lickliter, J.D., McGlave, P.B., DeFor, T.E., Miller, J.S., Ramsay, N.K., Verfaillie, C.M., Burns, L.J., Wagner, J.E., Eastlund, T., Dusenbery, K., and Weisdorf, D.J. (2000). Matched-pair analysis of peripheral blood stem cells compared to marrow for allogeneic transplantation. Bone Marrow Transplant. 26, 723–728.
- Lin, S.J., Yang, M.H., Chao, H.C., Kuo, M.L., and Huang, J.L. (2000). Effect of interleukin-15 and Flt3-ligand on natural killer cell expansion and activation: umbilical cord vs. adult peripheral blood mononuclear cells. Pediatr. Allergy Immunol. 11, 168–174.
- MacKey, M.C. (2001). Cell kinetic status of haematopoietic stem cells. Cell. Prolif. 34, 71–83.
- Marshak, D.R., Gottlieb, D., Kiger, A.A., Fuller, M.T., Kunath, T., Hogan, B., Gardner, R.L., Smith, A., Klar, A.J.S., Henrique, D., D'Urso, G., Datta, S., Holliday, R., Astle, C.M., Chen, J., Harrison, D.E., Xie, T., Spradling, A., Andrews, P.W., Przyborski, S.A., Thomson, J.A., Kunath, T., Strumpf, D., Rossant, J., Tanaka, S., Orkin, S.H., Melchers, F., Rolink, A., Keller, G., Pittenger, M.F., Marshak, D.R., Flake, A.W., Panicker, M.M., Rao, M., Watt, F.M., Grompe, M., Finegold, M.J., Kritzik, M.R., Sarvetnick, N., and Winton, D.J. (2001). Stem cell biology, Marshak, D.R., Gardner, R.L., and Gottlieb, D. eds. (Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press).
- McSweeney, P.A. and Storb, R. (1999). Mixed chimerism: preclinical studies and clinical applications. Biol. Blood Marrow Transplant. 5, 192–203.
- Medvinsky, A. and Dzierzak, E. (1996). Definitive hematopoiesis is autonomously initiated by the AGM region. Cell. 86, 897–906.
- Moore, K.A., Ema, H., and Lemischka, I.R. (1997). In vitro maintenance of highly purified, transplantable hematopoietic stem cells. Blood. 89, 4337–4347.
- Morrison, S.J., Prowse, K.R., Ho, P., and Weissman, I.L. (1996). Telomerase activity in hematopoietic cells is associated with self- renewal potential. Immunity. 5, 207–216.
- Negrin, R.S., Atkinson, K., Leemhuis, T., Hanania, E., Juttner, C., Tierney, K., Hu, W.W., Johnston, L.J., Shizurn, J.A., Stockerl-Goldstein, K.E., Blume, K.G., Weissman, I.L., Bower, S., Baynes, R., Dansey, R., Karanes, C., Peters, W., and Klein, J. (2000). Transplantation of highly purified CD34+ Thy-1+ hematopoietic stem cells in patients with metastatic breast cancer. Biol. Blood Marrow Transplant. 6, 262–271.
- Oh, I.H., Lau, A., and Eaves, C.J. (2000). During ontogeny primitive (CD34+ CD38-) hematopoietic cells show altered expression of a subset of genes associated with early cytokine and differentiation responses of their adult counterparts. Blood. 96, 4160–4168.
- Orlic, D., Girard, L.J., Anderson, S.M., Pyle, L.C., Yoder, M.C., Broxmeyer, H.E., and Bodine, D.M. (1998). Identification of human and mouse hematopoietic stem cell populations expressing high levels of mRNA encoding retrovirus receptors. Blood. 91, 3247–3254.
- Orlic, D., Bock, T.A., and Kanz, L. (1999). Hematopoietic stem cells biology and transplantation. Annals of The New York Academy of Sciences (New York, NY).
- Perkins, A.C. (1998). Enrichment of blood from embryonic stem cells in vitro. Reprod. Fertil. Dev. 10, 563–572.
- Pittenger, M.F., Mackay, A.M., Beck, S.C., Jaiswal, R.K., Douglas, R., Mosca, J.D., Moorman, M.A., Simonetti, D.W., Craig, S., and Marshak, D.R. (1999). Multilineage potential of adult human mesenchymal stem cells. Science. 284, 143–147.
- Shamblott, M.J., Axelman, J., Littlefield, J.W., Blumenthal, P.D., Huggins, G.R., Cui, Y., Cheng, L., and Gearhart, J.D. (2001). Human embryonic germ cell derivatives express a broad range of developmentally distinct markers and proliferate extensively in vitro. Proc. Natl. Acad. Sci. U. S. A. 98, 113–118.
- Sharp, J.G., Kessinger, A., Lynch, J.C., Pavletic, Z.S., and Joshi, S.S. (2000). Blood stem cell transplantation: factors influencing cellular immunological reconstitution. J. Hematother. Stem Cell Res. 9, 971–981.
- Smith, L.G., Weissman, I.L., and Heimfeld, S. (1991). Clonal analysis of hematopoietic stem-cell differentiation in vivo. Proc. Natl. Acad. Sci. U. S. A. 88, 2788–2792.
- Spangrude, G.J., Heimfeld, S., and Weissman, I.L. (1988). Purification and characterization of mouse hematopoietic stem cells. Science. 241, 58–62.
- Sudo, K., Ema, H., Morita, Y., and Nakauchi, H. (2000). Age-associated characteristics of murine hematopoietic stem cells. J. Exp. Med. 192, 1273–1280.
- Surbek, D.V., Steinmann, C., Burk, M., Hahn, S., Tichelli, A., and Holzgreve, W. (2000). Developmental changes in adhesion molecule expressions in umbilical cord blood CD34+ hematopoietic progenitor and stem cells. Am. J. Obstet. Gynecol. 183, 1152–1157.
- Tavian, M., Coulombel, L., Luton, D., Clemente, H.S., Dieterlen-Lievre, F., and Peault, B. (1996). Aorta-associated CD34+ hematopoietic cells in the early human embryo. Blood. 87, 67–72.
- Tavian, M., Hallais, M.F., and Peault, B. (1999). Emergence of intraembryonic hematopoietic precursors in the pre-liver human embryo. Development. 126, 793–803.
- Thomas, E.D. and Clift, R.A. (1999). Allogenic transplantation for chronic myeloid leukemia. Thomas, E.D., Blume, K.G., and Forman, S.J. eds. Blackwell Sci., 807–815.
- Till, J.E. and McCullough, E.A. (1961). A direct measurement of the radiation sensitivity of normal mouse bone marrow cells. Radiat. Res. 14, 213–222.
- U.S. Department of Health and Human Services. (2000). Report to Congress on the Status of Umbilical Cord Blood Transplantation.
- Van Zant, G., Rummel, S.A., Koller, M.R., Larson, D.B., Drubachevsky, I., Palsson, M., and Emerson, S.G. (1994). Expansion in bioreactors of human progenitor populations from cord blood and mobilized peripheral blood. Blood Cells. 20, 482–490.
- Verfaillie, C. M., personal communication.
- Verlinden, S.F., Mulder, A.H., de Leeuw, J.P., and van Bekkum, D.W. (1998). T lymphocytes determine the development of xeno GVHD and of human hemopoiesis in NOD/SCID mice following human umbilical cord blood transplantation. Stem Cells. 16, Suppl. 1, 205–217.
- Wang, M.W., Consoli, U., Lane, C.M., Durett, A., Lauppe, M.J., Champlin, R., Andreeff, M., and Deisseroth, A.B. (1998). Rescue from apoptosis in early (CD34-selected) versus late (non-CD34-selected) human hematopoietic cells by very late antigen 4- and vascular cell adhesion molecule (VCAM) 1-dependent adhesion to bone marrow stromal cells. Cell Growth Differ. 9, 105–112.
- Weissman, I. L., personal communication.
- Whetton, A.D. and Graham, G.J. (1999). Homing and mobilization in the stem cell niche. Trends Cell. Biol. 9, 233–238.
- Wright, D.E., Cheshier, S.H., Wagers, A.J., Randall, T.D., Christensen, J.L., and Weissman, I.L. (2001). Cyclophosphamide/granulocyte colony-stimulating factor causes selective mobilization of bone marrow hematopoietic stem cells into the blood after M-phase of the cell cycle.
- Zon, L. I., personal communication.
 Chapter 4 | Table of Contents | Chapter 6
Chapter 4 | Table of Contents | Chapter 6 
Historical content: June 17, 2001