
Maps of Distribution and Abundance of Selected Species of Birds on Uncultivated Native Upland Grasslands and Shrubsteppe in the Northern Great Plains
Harold A. Kantrud and James E. Roelle
Many areas of the northern Great Plains (Fenneman 1931) remain poorly known ornithologically. Knowledge of species distribution in portions of this vast area has been advanced by recent State birds books (Stewart 1975; South Dakota Ornithologists' Union 1978) but many voids remain. Especially desirable for biologists and land managers is information concerning the relative abundance of plant and animal species over large land areas. This information can only be obtained from extensive surveys of the biota. The field of community ecology has been severely hampered by lack of information on species abundance (Dice 1930; Cain 1947; Partch 1962; Udvardy 1963).
In 1974-78, the U.S. Fish and Wildlife Service's Migratory Birds and Biological Services Programs jointly funded an extensive study in an attempt to develop a classification of native, northern Great Plains grasslands based on their use by breeding birds. Other goals were to investigate the effects of grazing and other environmental factors on breeding birds and the vegetation. The results of these portions of the study will be reported later.
The purpose of this report is to provide ecologists and ornithologists with recent information on areas of abundance of some of the more important members of the avifauna which use grasslands. This information should also prove valuable to conservationists and resource managers as energy development and further expansion and intensification of the agricultural industry result in increased destruction of native grassland habitat in areas now supporting some of the less common species.
During the study, breeding bird censuses were conducted on 615 plots of uncultivated native upland grassland and shrubsteppe in the portion of the northern Great Plains that lies within the United States (Fig. 1). The study area (about 6 X 103 km2) includes portions of Montana, Wyoming, Colorado, North Dakota, South Dakota, and Nebraska. Grasslands in the mountains, badlands, sand deposits, and river valleys (shown in black on Fig. 1) were not sampled and so are excluded from further consideration.
Contents
Figures
- Figure 1--Outline of Northern Great Plains Study Area
- Figure 2--Location of Soils and Sample Plots
- Figure 3--Quarter Section Sample Plot and Transect Route
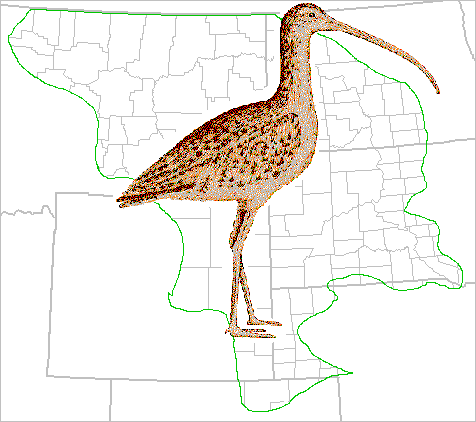
- Figure 4--Breeding Distribution of the Long-billed Curlew (Numenius americanus)
- Figure 5--Breeding Distribution of the Upland Sandpiper (Bartramia longicauda)
- Figure 6--Breeding Distribution of the Horned Lark (Eremophila alpestris)
- Figure 7--Breeding Distribution of the Sage Thrasher (Oreoscoptes montanus)
- Figure 8--Breeding Distribution of the Sprague's Pipit (Anthus spragueii)
- Figure 9--Breeding Distribution of the Common Yellowthroat (Geothlypis trichas)
- Figure 10--Breeding Distribution of the Bobolink (Dolichonyx oryzivorus)
- Figure 11--Breeding Distribution of the Western Meadowlark (Sturnella neglecta)
- Figure 12--Breeding Distribution of the Red-winged Blackbird (Agelaius phoeniceus)
- Figure 13--Breeding Distribution of the Brewer's Blackbird (Euphagus cyanocephalus)
- Figure 14--Breeding Distribution of the Brown-headed Cowbird (Molothrus ater)
- Figure 15--Breeding Distribution of the Lark Bunting (Calamospiza melanocorys)
- Figure 16--Breeding Distribution of the Savannah Sparrow (Passerculus sandwichensis)
- Figure 17--Breeding Distribution of the Grasshopper Sparrow (Ammodramus savannarum)
- Figure 18--Breeding Distribution of the Baird's Sparrow (Ammodramus bairdee)
- Figure 19--Breeding Distribution of the Vesper Sparrow (Pooecetes grmaineus)
- Figure 20--Breeding Distribution of the Clay-colored Sparrow (Spizella pallida)
- Figure 21--Breeding Distribution of the Brewer's Sparrow (Spizella breweri)
- Figure 22--Breeding Distribution of the McCown's Longspur (Calcarius mccownii)
- Figure 23--Breeding Distribution of the Chestnut-collared Longspur (Calcarius ornatus)
Methods
SELECTION OF SAMPLE UNITSSeveral different methods were used to select the sample units (Fig. 2) over the years. In 1974, plots were selected in numbers proportional to the sizes of the areas of the several mapped physiographic landforms in North Dakota (U.S. and North Dakota Geological Survey maps). Sample unit locations were initially selected at random from numbered legal townships, sections, and quarter sections. However, differential rates of agricultural disturbance of grasslands among the landforms prevented maintenance of randomness. If a selected area was devoid of native grassland, the nearest usable plot was chosen. Many plots were rejected after field examination revealed old furrows, rockpiles, domestic grass plantings, or other evidence of past cultivation. Although most of the plots studied in 1974 were legal quarter sections (65 ha), several were as large as a section or as small as 16 ha. Most plots were square but occasionally it was necessary to utilize odd-shaped areas.
In 1975, in Montana and Wyoming, sample units were randomly selected from consecutively numbered townships, in proportion to the area of grassland types mapped by Kuchler (1964). After a township was selected and found to lie entirely within the Kuchler type, one out of a set of 27 possible clusters of four quarter sections within the township was selected at random. In the field, the usable quarter section nearest to each of the four cluster elements was censused.
In 1976, in Colorado, Montana, Nebraska, and South Dakota, single quarter section sample units were selected in proportion to the areas of grassland types 64, 65, and 66 of Kuchler (1964). Acceptable plots nearest to the southwest corner of every eighth township in South Dakota and every fourth township in the other States were censused.
In 1977-78, the entire study area was sampled. Quarter section sample units were subjectively chosen in numbers proportional to the areas of major soil groups mapped by Aandahl (1972).
Throughout the study, grasslands in mountains, badlands, sand deposits, or river valleys were not sampled ( Fig. 1). On a local scale, grasslands not considered typical of general area soils were also rejected. These included expanses of wet meadow and rocky hilltops, dune sands, and badlands. The major soil groups of Aandahl (1972) were later used to stratify all the sample plots ( Fig. 2). A description of these soils and the birds and vegetation associated with them is reported elsewhere (Kantrud and Kologiski 1982).
BIRD CENSUSES
A single bird census was conducted on each plot at times between sunrise and sunset when sustained wind velocities did not exceed 24 km/h, temperatures did not exceed 32°C, and precipitation did not occur. Field work was conducted by two teams, each consisting of a botanist and an ornithologist. Census dates were as follows: 1974--22 May to 19 July; 1975--29 May to 18 July; 1976--23 May to 14 July; 1977--24 May to 13 July; 1978--22 May to 2 July.
Comparison of five 3-h time periods (0600-2100) during this study showed that: (1) of 29 species censused, peak densities for one or more species occurred during each of the five periods; (2) peak densities for 11 species occurred during the period 0900 to 1200 hours; and (3) observed total density decreased by up to 25% during counts conducted after the period 0600 to 0900 hours. Temporal variation in bird counts undoubtedly biased density indices for some species but was not considered a serious problem because of its random nature as applied to the censuses.
Analyses based on single bird censuses are not recommended during intensive studies where population estimates are required. Nevertheless, they have been used successfully to derive indices to populations or multispecies diversity measurements during extensive surveys (Robbins and Van Velzen 1967; Stewart and Kantrud 1972; United States Department of the Interior and Canadian Department of the Environment 1977; Rotenberry 1978; Kantrud 1981). Speirs and Orenstein (1967) have showed that in open Ontario habitats, average efficiency of single censuses is only 66-76% that of 6-10 censuses in estimating whole populations. However, 1 felt justified in basing indices of bird density on single counts in the grassland habitats because most of the species have behavioral adaptations--including lengthy, loud, and often elaborate flight songs, perch songs, and synchronous display--that tend to increase greatly their detectability compared with birds inhabiting wooded areas. Furthermore, birds are rather easily flushed in grasslands. In addition, the large size of the sample plots resulted in relatively high absolute numbers of bird observations per plot.
The teams attempted to compensate for phenological differences across the northern Great Plains by beginning the surveys in southerly areas of low elevation. As the breeding season progressed, the surveys were moved to regions of successively higher latitude and elevation.
All species were identified in the field by sight or sound, or both. Counts were based primarily on number of pairs as indicated by territorial males or segregated pairs. Each male blackbird or brown-headed cowbird (Molothrus ater) was considered to represent a pair. Single shorebirds observed on a sample plot were considered to represent a pair; if more than one was observed, the total was halved and rounded to the nearest lower integer. Additional details on determinations of breeding status were described by Stewart and Kantrud (1972).
The "hollow square" technique of Speirs and Orenstein (1967) was used on the standard 65-ha tracts. The 1609-m (1-mile) transect route lay 201 m inside the boundary ( Fig. 3). Birds were counted within 101 m of the route, yielding an areal coverage of 31.5 ha. Transect routes were variable on the few tracts of other sizes and shapes. The verges of many plots bordered other habitat types, so to reduce the influence on the counts of birds occupying fencelines or adjacent habitats, census transects were restricted to the interior of each tract except in the few tracts < 65 ha.
Transects were walked at a standard speed of 40 m/min. To allow standardization of data from plots or transects of unequal size and shape, expression of bird density in units of time has been used in similar extensive studies (e.g., Grinnell and Storer 1924; Skinner 1975). So, the indices to bird density were expressed in pairs/min X 103.
GRAZING INTENSITY
Grazing intensity was estimated by comparing the percentage of bare soil, amount of standing and fallen dead vegetation, and average vegetative height on the study plots with that of nearby protected or ungrazed areas. On the basis of this information, plots were placed into the three subjective categories of heavily, moderately, and lightly grazed. Distribution of the 615 plots among grazing categories was as follows: heavy, 197; moderate, 241; and light, 177. Any plots lacking evidence of livestock use were not censused. Most of the plots were grazed by cattle but a few were used for sheep, bison, or horses. A few plots contained various combinations of domestic livestock and horses. Plots in each grazing category were well distributed among the strata.
Results
Maps (Figs. 4-23) are provided for 20 of the 29 species studied. Species not mapped include those with too few records to establish a meaningful distribution (ferruginous hawk, Buteo regalis; northern harrier, Circus cyaneus; sharp-tailed grouse, Pedioecetes phasianellus; sage grouse, Centrocercus urophasianus; killdeer, Charadrius vociferus; mountain plover, Charadrius montanus; mourning dove, Zenaida macroura; burrowing owl, Athene cunicularia; common nighthawk, Chordeiles minor).
Some mapped species achieve greater abundance and wider distribution in wetland or other habitats but have been included to show their restricted distribution on upland native grasslands in the northern Great Plains. Four colors of map symbols are used to delineate areas of abundance and distribution for each species on the habitat studied. The symbols represent units of abundance based on division of the density indices for each species into the upper (black dot), middle (blue dot), and lower (red dot) tercile groups. Terciles are defined as points dividing the distribution of observed species density (pairs/min X 103) values among the sample units into thirds. The green map symbols (green dots) represent breeding bird locations taken from supplementary counts and incidental observations gathered on tracts of native grassland and shrubsteppe encountered during travel between sample units. Bird nomenclature is according to American Ornithologists' Union (1957) and its several supplements.
Discussion and Conclusions
Distribution maps normally depict a species' range regardless of the variety of habitats used. Bird records mapped here were obtained within the single, broad habitat of grazed upland native grassland and shrubsteppe. Distributions thus derived must not be interpreted as overall distributions of the species within the study area, although the two distributions would probably be very similar for some of the "pure" grassland birds.
The habitat investigated included various types of shortgrass and mixedgrass prairie with or without short (< 1 m) shrubs, and shrubsteppe dominated by mixtures of shrubs and grasses of mid and short height. Each of these types is attractive to certain bird species. The maps may also reflect the variable effects of grazing intensity on distribution and abundance of individual species in different areas of the region studied. In addition, extensive cultivation of grasslands for agriculture may have reduced the breeding distribution of some species that require large foraging areas. Finally, many of the species have their center of abundance outside the northern Great Plains region. Breeding populations of these species extend various distances into the region from different directions.
Common yellowthroat (Geothlypis trichas) (Fig. 9), bobolink (Dolichonyx oryzivorus) (Fig. 10), red-winged blackbird (Agelaius phoeniceus) (Fig. 12), and savannah sparrow (Passerculus sandwichensis) (Fig. 16) are not typical of the dry upland sites studied. All but the latter had rather small (< 1/2 the study area) distribution along the eastern border of the study area. Breeding populations of these species are normally found in dense wetland, meadow, or tallgrass vegetation. The limit of their ability to breed in dryer upland sites is likely reached in the more mesic mixed grass prairie along the eastern edge of the northern Great Plains. The disjunct distribution of the savannah sparrow (Fig. 16) indicates that it is of fairly regular occurrence in the cooler, mesic grasslands of central North Dakota and northeastern Montana but becomes rare as a breeding bird in adjacent arid grasslands to the west. However, this sparrow reappears in abundance in the northwestern portion of the study area in mesic foothills grasslands.
The remaining 16 species are more or less typical of dry upland habitats. The commonest 10 species are extensively (> 1/2 the study area) distributed. Of these 10 species, upland sandpiper (Bartramia longicauda) (Fig. 5), brown-headed cowbird (Fig. 14), and grasshopper sparrow (Ammodramus savannarum) (Fig. 17) reach greatest abundance in the eastern mixed grass prairie. High populations of chestnut-collared longspur (Calcarius ornatus) (Fig. 23) also occur in this area and extend across northern Montana. Brewer's blackbird (Euphagus cyanocephalus) (Fig. 13), vesper sparrow (Pooecetes gramineus) (Fig. 19), and Brewer's sparrow (Spizella breweri) (Fig. 21) reach greatest abundance in the shrubsteppe zone of northeastern Wyoming and southeastern Montana. The horned lark (Eremophila alpestris) (Fig. 6) likely has three areas of peak abundance. These include shortgrass prairie near the intersection of the Colorado-Nebraska-Wyoming borders, an area in east-central Montana, and in the part of Montana that formed the northwestern corner of the study area. The western meadowlark (Sturnella neglecta) (Fig.11) probably reaches peak densities in southeastern Montana, but high populations also occur regularly eastward through southern North Dakota and most of South Dakota. The lark bunting (Calamospiza melanocorys) (Fig. 15) seemingly has an unusual pattern of abundance in the northern Great Plains. High populations occur in a zone roughly extending from southeastern South Dakota to central Montana, then southerly through the shrubsteppe area of Montana and Wyoming into the shortgrass area of northeastern Colorado and the southwestern portion of the Nebraska panhandle. For unknown reasons, populations appear sparse in a zone extending northwestward from northwestern Nebraska to the southwest corner of Montana.
Six species typical of the dry upland sites studied likely have small or disjunct distributions. The long-billed curlew (Numenius americanus) (Fig. 4), a formerly widespread species in the study area, has disjunct populations in western South Dakota and the northcentral Montana grasslands. The northern population probably is N. a. parvus and the southern N. a. americanus. Observations indicate that the foothills grasslands of Montana harbor the largest remaining populations. The sage thrasher (Oreoscoptes montanus) (Fig. 7) is restricted to the shrubsteppe area of Wyoming and southeastern Montana. The Sprague's pipit (Anthus spragueii) (Fig. 8) reaches greatest density from northwestern North Dakota across northern Montana; high populations also occur in some of the Montana foothills grassland. Baird's sparrow (Ammodramus bairdii) (Fig. 18) has a distribution very similar to that of Sprague's pipit and upland nesting populations of savannah sparrow. Disjunct areas of greatest abundance occur in the northwestern North Dakota-northeastern Montana regions and in the northwestern corner of the study area. Dense populations of clay-colored sparrow (Spizella pallida) (Fig. 20) are found in the northeastern edge of the study area on grasslands that contain dense stands of wolfberry buckbrush (Symphoricarpos occidentalis). Disjunct areas of abundance are indicated for McCown's longspur (Calcarius mccownii) (Fig. 22). These include the shortgrass area of southeastern Wyoming and northeastern Colorado, small areas of shortgrass and birdsfoot sage (Artemisia pedatifida) in the area of Wyoming generally dominated by A. tridentata shrubsteppe, and the two areas of Montana described for the horned lark.
Acknowledgments
R. E. Stewart supervised and participated in the field work during 1974-75. Biological assistance was received from B. A. Barnes, C. A. Faanes, B. A. Hanson, R. L. Kologiski, and J. T. Lokemoen. D. A. Davenport, A. M. Frank, and D. H. Johnson were responsible for computer programming and statistical analyses. The study was supported by the U.S. Fish and Wildlife Service through the Migratory Birds Program and the Coal Project, Western Energy and Land Use Team, Office of Biological Services.
Literature Cited
Aandahl, A. R. 1972. Soils of the Great Plains. [Map]. A. R. Aandahl, P.O Box 81242, Lincoln, Nebraska 68508.
American Ornithologists' Union. 1957. Check-list of North American birds. 5th ed. and suppl. Port City Press, Baltimore. 691 pp.
Cain, S. A. 1947. Characteristics of natural areas and factors in their development. Ecol. Monogr. 17:185-200.
Dice, L. R. 1930. Methods of indicating relative abundance of birds. Auk 47:22-24.
Fenneman, N. M. 1931. Physiography of western United States. McGraw-Hill Co., New York. 534 pp.
Grinnell, J., and T. I. Storer. 1924. Animal life in the Yosemite. University of California Press, Berkeley. 752 pp.
Kantrud, H. A. 1981. Grazing intensity effects on the breeding avifauna of North Dakota native grasslands. Can. Field-Nat. 95:404-417.
Kantrud, H. A., and R. L. Kologiski. 1982. Effects of soils and grazing on breeding birds of uncultivated upland grasslands of the northern Great Plains. U.S. Fish Wildl. Serv. Wildl. Res. Rept. 15.
Kuchler, A. W. 1964. Potential natural vegetation of the conterminous United States. American Geographical Society, New York. 39 pp. + 116 plates.
Partch, M. L. 1962. Species distribution in a prairie in relation to waterholding capacity. Proc. Minn. Acad. Sci. 31:38-43.
Robbins, C. S., and W. T. Van Velzen. 1967. The breeding bird survey, 1966. U.S. Fish Wildl. Serv. Spec. Sci. Rep. Wildl. 102. 43 pp.
Rotenberry, J. T. 1978. Components of avian diversity along a multifactorial climatic gradient. Ecology 59:693-699.
Skinner, R. M. 1975. Grassland use patterns and prairie bird populations in Missouri. Pages 171-180 in M. K. Wali, ed. Prairie: A multiple view. University of North Dakota Press, Grand Forks.
South Dakota Ornithologists' Union. 1978. Birds of South Dakota. W. H. Over Museum, Vermillion. 311 pp.
Speirs, J. M., and R. Orenstein. 1967. Bird populatfons in fields of Ontario County, 1965. Can. Field-Nat. 81:175-183.
Stewart, R. E. 1975. Breeding birds of North Dakota. Tri-college Center for Environmental Studies, Fargo. 295 pp.
Stewart, R. E., and H. A. Kantrud. 1972. Population estimates of breeding birds in North Dakota. Auk 89:766-788.
Udvardy, M. D. F. 1963. Bird faunas of North America. Proc. Int. Ornithol. Congr. 13:1147-1167.
United States Department of the Interior and Canadian Department of the Environment. 1977. Standard operating procedures for aerial waterfowl breeding ground population and habitat surveys. Section IV. 33 pp.
This resource is based on the following source (Northern Prairie Publication 0517):
Kantrud, Harold A. 1982. Maps of distribution and abundance of selected species of birds on uncultivated native upland grasslands and shrubsteppe in the Northern Great Plains. U.S. Department of the Interior, Fish and Wildlife Service, FWS/OBS-82/31. 31pp.
This resource should be cited as:
Kantrud, Harold A. 1982. Maps of distribution and abundance of selected species of birds on uncultivated native upland grasslands and shrubsteppe in the Northern Great Plains. U.S. Department of the Interior, Fish and Wildlife Service, FWS/OBS-82/31. Jamestown, ND: Northern Prairie Wildlife Research Center Online. http://www.npwrc.usgs.gov/resource/birds/plainmap/index.htm (Version 16JUL97).
This report was performed for:
Western Energy and Land Use Team
Office of Biological Services
Fish and Wildlife Service
U.S. Dept. of the Interior
Washington, D.C. 20240and
Migratory Birds Program
Fish and Wildlife Service
U.S. Dept. of the Interior
Washington, D.C. 20240
Downloading Instructions -- Instructions on downloading and extracting files from this site.
Installation: Extract all files and open index.htm in a web browser.plainmap.zip ( 366K ) -- Maps of Distribution and Abundance of Selected Species of Birds on Uncultivated Native Upland Grasslands and Shrubsteppe in the Northern Great Plains
