| |
 |
|
Developmental Alcohol and Circadian Clock Function David
J. Earnest, Ph.D., Wei-Jung A. Chen, Ph.D., DAVID J. EARNEST, PH.D., is an associate professor, WEI-JUNG A. CHEN, PH.D., is an assistant professor, and JAMES R. WEST, PH.D., is a professor in the Department of Human Anatomy and Medical Neurobiology, Texas A&M University Health Science Center, College of Medicine, College Station, Texas. |
Studies
in rats found that alcohol exposure during the early postnatal period, particularly
during the brain-growth-spurt period, can result in cell loss in various brain
regions and persistent behavioral impairments. Some investigators have speculated
that the body’s internal clock, which is located in the suprachiasmatic
nuclei (SCN) in the brain, may also be affected by developmental alcohol exposure.
For example, alcohol-induced damage to the SCN cells and their function could
result in disturbances of the circadian timekeeping function, and these disturbances
might contribute to the behavioral impairments and affective disorders observed
in people prenatally exposed to alcohol. Preliminary findings of studies conducted
in rats suggest that developmental alcohol exposure may indeed interfere with
circadian clock function as evidenced by a shortened circadian sleep-wake
cycle and changes in the release of certain brain chemicals (i.e., neuropeptides)
by SCN cells. KEY
WORDS: circadian rhythm; sleep disorder; fetal alcohol effects; prenatal alcohol
exposure; fetal alcohol syndrome; neuropeptides; alcohol-related neurodevelopmental
disorder; cytolysis; nerve growth factors; CNS nuclei
Since ancient Roman times, people have suspected that maternal alcohol consumption has deleterious effects on the developing fetus. The clinical diagnosis of fetal alcohol syndrome (FAS), however, was not defined until 1973 when Jones and Smith first documented a constellation of characteristics in the offspring of mothers who had abused alcohol during pregnancy. These characteristics include a set of unique facial features, such as a flat groove between the nose and upper lip (i.e., a flat philtrum), a thin upper lip, and a small nose, as well as central nervous system (CNS) dysfunction. The CNS defects--which include reduced brain size (i.e., microencephaly), reductions in the volume of various brain regions (e.g., in the anterior cerebellum1 (1 The anterior cerebellum is involved in motor coordination and maintenance of posture.) and the corpus callosum2) (2 The corpus callosum is a bundle of nerve fibers that connects the two cerebral hemispheres and mediates information transfer between those hemispheres.) and behavioral impairments (e.g., learning difficulties and attention deficits) (Roebuck et al. 1999)--are by far the most debilitating features associated with FAS (Stratton et al. 1996).
During the past 20 years, studies using animal model systems have generated substantial information on the risk factors associated with maternal alcohol use and the spectrum of alcohol-induced brain injuries among affected offspring. Perhaps most importantly, these animal studies have identified the brain-growth-spurt period as the developmental stage most vulnerable to alcohol-induced brain injury. In humans, this particular brain-growth period begins during the third trimester of pregnancy, peaks around birth, and persists through the first few years of life. In rats, which are the most commonly used species in fetal alcohol research, the corresponding rapid brain-growth period occurs during the first 2 weeks after birth.
Alcohol
exposure in rats during this early postnatal period produces structural changes
in the brain comparable to those observed in human FAS patients. These changes
range from reductions in the rat’s brain size and weight (Maier et al.
1996) to the loss of various cell populations in discrete brain regions, such
as the cerebellum and the hippocampus3 (
One key question in the fetal alcohol research field is whether developmental alcohol exposure has long-term consequences. In humans, the characteristic facial abnormalities associated with FAS typically become less prominent with age. However, many reported findings suggest that alcohol-induced deficits in brain development may persist into adulthood. Using noninvasive techniques (e.g., magnetic resonance imaging and positron emission tomography) in a small number of children with FAS, researchers have confirmed brain damage from prenatal alcohol exposure and have determined the related behavioral consequences, including learning deficits, hyperactivity, lack of inhibition, depression, and sleep disturbances (Hannigan et al. 1995; Roebuck et al. 1999).
Studies using animal models have been invaluable in establishing the association between structural brain damage and long-term behavioral impairments. For example, researchers have linked the cell loss that occurs in a rat’s cerebellum and hippocampus from early postnatal alcohol exposure with deficits in motor performance (Thomas et al. 1996) and in learning and memory (Goodlett et al. 1987), respectively. These experimental data could provide a focus for long-term clinical investigations in humans, because some of the FAS patients identified and followed since 1973 have now reached adulthood. Thus, researchers could use noninvasive techniques to determine potential structural brain damage in FAS patients and correlate the findings with adult behavioral impairments.
A recent
and still highly speculative area of research focuses on whether prenatal
alcohol exposure also has long-term effects on the regulation of the body’s
internal clock and whether these potential effects could contribute to some
of the previously mentioned behavioral problems associated with FAS. This
article first provides a brief review of the structure and function of this
internal clock. It then discusses the potential relationship between prenatal
alcohol exposure and disturbances in the internal clock, including preliminary
results of animal studies assessing the effects of developmental alcohol exposure
on circadian rhythms.
The
Structure and Function of the Internal Biological Clock
In mammals,
a variety of biological processes, such as the levels of various brain chemicals
(i.e., neurotransmitters), blood hormone concentrations, and sleep-wake behavior,
undergo rhythmic fluctuations that closely parallel the time course of the
daily solar cycle. This temporal organization of mammalian biology to approximate
the 24-hour cycle of change in the physical environment occurs even in the
absence of external time cues; consequently, these rhythms have been termed
circadian. In mammals, including humans, the internal biological clock is
responsible for the generation of all circadian rhythms and their synchronization
(i.e., entrainment) to light-dark cycles. During entrainment, mammalian circadian
rhythms assume not only the periodicity of the light-dark cycle, but also
adopt a specific phase relationship to this environmental cycle. Structurally,
the internal clock is discretely localized within the brain in paired clusters
of nerve cells (i.e., neurons) called the suprachiasmatic nuclei (SCN). Consistent
with their circadian timekeeping function, these cells rhythmically release
certain brain chemicals (i.e., neuropeptides), such as brain-derived neurotrophic
factor (BDNF) and arginine vasopressin (AVP). These neuropeptides as well
as other peptides (e.g., vasoactive intestinal polypeptide [VIP]), in turn,
may act on other brain regions or peripheral organs to generate a variety
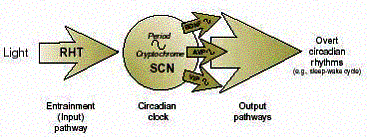
of overt circadian rhythms, such as the sleep-wake cycle (see figure).
|
|
|
Figure: Functional components of the internal timekeeping system responsible for the regulation of circadian, or 24-hour, rhythms in mammals. The central circadian pacemaker, or “clock,” is located in the brain in the suprachiasmatic nuclei (SCN). These cells contain the “gears” that are necessary for accurate timekeeping. These gears consist of certain genes (e.g., the Period genes and the Cryptochrome genes) that are rhythmically expressed and regulate ciracadian output pathways. These path-ways involve various signals that emanate from the SCN, including neuropeptides, such as brain-derived neurotrophic factor (BDNF), arginine vasopressin (AVP), and vasoactive intestinal polypeptide (VIP). These neuropeptides and other output signals act on other brain regions or peripheral organs and result in the generation of overt rhythms in biochemical, physiological, and behavioral processes throughout the body. The retinohypothalamic tract (RHT) is an entrainment, or input, pathway that mediates the synchronization of the circadian clock to the light-dark cycle. |
The normal circadian timekeeping function of the SCN is crucial for maintaining human health and performance by providing for the temporal coordination of internal physiological processes with each other and with the daily light-dark cycle. Damage to the SCN, which would compromise this temporal organization, could thus affect the body’s susceptibility to physiological disorders.
To date, the neurobehavioral consequences of many health problems, including developmental alcohol exposure, on the regulation of circadian rhythmicity are virtually unknown. In animal models, complete destruction of the SCN is necessary to abolish circadian rhythmicity, although experimental or natural injury to subgroups of SCN neurons can produce tangible changes in the SCN-mediated regulation of overt physiological and behavioral rhythms. For example, in rats, partial damage to the SCN alters the animals’ circadian activity rhythm and its pattern of entrainment to light-dark cycles (Pickard and Turek 1985). 4 (4 In nocturnal rodents, activity levels typically rise near the onset of the dark phase of the daily cycle, whereas rest periods occur primarily during the light phase.) Specifically, the destruction of some, but not all, SCN cells results in daily onset of activity at earlier times than normal during exposure to constant environmental conditions and during synchronization to a light-dark cycle.
In a similar
fashion, the aging process has been shown to affect discrete components of
the human and rodent SCN and the brain chemicals released by the SCN (Roozendaal
et al. 1987; Swaab et al. 1988; Krajnak et al. 1998). This impact of aging
on SCN cellular organization is associated with functional disturbances in
the generation and regulation of circadian behavior, which is often reflected
in the sleep-wake cycles of both humans and rodents (Ingram et al. 1982; Moore
1991). For example, aging-related changes can occur in many properties of
sleep-wake rhythmicity, including circadian period, entrainment to light-dark
cycles, and resetting or phase-shifting to earlier or later times by light
(Pittendrigh and Daan 1974; Schwartz 1993), although the reported effects
and their relative magnitudes differ considerably between studies. The amplitude
of the sleep-wake rhythm also decreases during aging such that day-night differences
in activity levels are diminished. Another change in sleep-wake rhythmicity
that occurs with advancing age is that sleep becomes more fragmented, resulting
in increased arousals and time awake during the night and in a reduction in
the total amount of deep sleep. In humans and rodents, these age-related perturbations
in the circadian regulation of behavior are accompanied by the partial loss
of various neuropeptide-releasing cell types in the SCN and by a damping of
the daily peak in SCN levels of these neuropeptides (Roozendaal et al. 1987;
Swaab et al. 1988; Krajnak et al. 1998).
Alcohol’s
Potential Influences on the Internal Clock
Based on the well-established evidence that alcohol exposure during rapid brain growth causes cell loss, alters information pathways between brain regions, and decreases the production of brain chemicals responsible for the communication among neurons, it is reasonable to speculate that alcohol may also adversely affect SCN development. Such alcohol-induced insults during the period of rapid brain growth could produce cell loss and/or structural alterations in the SCN that could result in subsequent disturbances of the circadian timekeeping function. Although it is unlikely that the damage to the SCN would be severe enough to cause complete loss of circadian rhythmicity, more subtle disturbances might occur. For example, it is possible that developmental alcohol exposure could lead to permanent reductions in rhythm amplitude, changes in circadian period, and modulation of the SCN clock’s responses to light signals. These potential effects of developmental alcohol exposure on SCN timekeeping function could account for some of the behavioral disturbances observed in FAS patients. For example, depression has been identified as a behavioral manifestation of FAS and circadian rhythm abnormalities have been associated with bipolar affective disorder (i.e., manic-depressive illness) (Moore 1991; Schwartz 1993).
Perturbations in the normal periodicity of circadian rhythms and their synchronization to light-dark signals, whether caused by developmental alcohol exposure or other factors, would be expected to affect chronotherapeutic phenomena in the pharmacologic treatment of diseases, because the efficacy and side effects of many drugs are known to depend on the time of administration in relation to normal body rhythmicity. 5 (5 For example, chemotherapy agents are usually more effective in treating cancer and are associated with reduced side effects when they are administered in a timed or circadian pattern.) Moreover, such perturbations could exacerbate sleep disturbances associated with normal aging and affective disorders, and impair performance in shift workers (Schwartz 1993). Therefore, people with FAS who suffered brain damage within the SCN as a result of developmental alcohol exposure could experience such problems throughout their adult life.
Developmental alcohol exposure could interfere with the circadian clock and its timekeeping function at two levels (see figure). First, it could damage the clock mechanism itself by affecting the structure or function of cells in the SCN. In this case, the changes in circadian function would reflect either a global loss of SCN cells or alterations in the rhythmic activation (i.e., expression) of specific genes that are essential “gears” of the clockworks, such as the Period and Cryptochrome genes, with-out which the clock does not work.
Second, developmental alcohol exposure could interfere only with the “hands” of the clock--that is, with the SCN output signals (e.g., the neuropeptides BDNF, AVP, and VIP) and with overt circadian rhythms throughout the body that are regulated by these signals (e.g., the sleep-wake cycle). It will be important to examine the SCN of adults with FAS for evidence of developmental alcohol-induced cell loss, perturbations in the expression of clock genes, and/or alterations in neuropeptide output signals in order to determine the levels at which develop-mental alcohol could affect circadian rhythms. Once more information becomes available on the severity of the effects of developmental alcohol exposure on circadian clock function and on the mechanism by which alcohol-induced circadian disturbances occur, it may be possible to develop therapies to treat specific rhythm abnormalities.
Studies in rats using alcohol exposure regimens that are equivalent to third-trimester alcohol exposure in humans are beginning to provide specific evidence for the impact of developmental alcohol exposure on SCN rhythmicity and circadian behavior as described in the following paragraph. Furthermore, alcohol exposure has been shown to decrease the levels of a class of growth factors called neurotrophins, which includes BDNF, in other regions of the developing rat brain (Maier et al. 1999). Because heavy maternal alcohol consumption during human pregnancy has been linked to disturbances in the off-spring’s sleep-wake patterns (Rosett et al. 1979; Stratton et al. 1996), these observations suggest that developmental alcohol exposure might disrupt the rhythmic activity of the SCN and neuropeptide release and interfere with circadian behavior patterns.
To further explore this hypothesis, Earnest and colleagues (1997) examined the long-term effects of early postnatal alcohol exposure on circadian neurotrophin expression in the SCN and on the rhythm of locomotor activity in rats. Preliminary findings demonstrate that adult rats exposed to alcohol during this critical period of brain development exhibited a shortened circadian sleep-wake cycle, so that their onset of activity occurred much earlier each day. In addition, the activity of animals exposed to developmental alcohol was more fragmented, with frequent alter-nation between short intervals of sleep and waking. This alcohol-induced change in circadian behavior was associated with marked damping of the BDNF rhythm in the SCN so that peak levels during the night were greatly reduced. Because BDNF has been implicated in the regulation of SCN clock responses to light (Liang et al. 2000), the disruption of BDNF rhythmicity induced by developmental alcohol exposure may also interfere with the light-mediated control of circadian rhythms. Based on these observations, further analysis is warranted to characterize fully the effects of developmental alcohol exposure on circadian timekeeping and their association with damage to the SCN clock or its associated neuropeptides.
Conclusions
Although the analysis of the relationship between developmental alcohol exposure and circadian rhythmicity is still highly speculative, this research may hold potential in the analysis and treatment of alcohol’s deleterious effect on the developing organism. For example, many behaviors are regulated by numerous brain areas, making it difficult to study alcohol’s effects on those behaviors. In contrast, the discrete localization of circadian clock function in the SCN provides a unique opportunity to identify specific alcohol-related neurobehavioral disturbances and examine their relation to the permanent effects of developmental alcohol exposure on other brain regions. Furthermore, information on the long-term behavioral consequences of FAS is still limited; many characteristics of FAS are not always fully evident in adulthood. Consequently, the identification of permanent alcohol-related alterations in circadian timekeeping may ultimately be useful as a long-term index or behavioral marker for FAS. Finally, research on how the SCN and its circadian function are affected by developmental alcohol exposure could yield important information on the basic mechanisms underlying alcohol-induced injury during rapid brain growth. Such information also could lead to new strategies in the treatment of known behavioral correlates of FAS, such as depression and sleep-wake abnormalities, that result from a disruption of the internal timing of body processes.
References
BONTHIUS, D.J., AND WEST, J.R. Alcohol-induced neuronal loss in developing rats: Increased brain damage with binge exposure. Alcoholism: Clinical and Experimental Research 14:107-118, 1990.
EARNEST, D.J.; MAHONEY, J.C.; CHEN, W.-J.A.; SOHRABJI, F.; AND WEST, J.R. Alcohol exposure during rapid brain development mimics the effects of aging on BDNF expression in the SCN and circadian timekeeping. Society of Neuroscience Abstracts 23:20, 1997.
GOODLETT, C.R.; KELLY, S.J.; AND WEST, J.R. Early postnatal alcohol exposure that produces high blood alcohol levels impairs development of spatial navigation learning. Psychobiology 15:64-74, 1987.
HANNIGAN, J.H.; MARTIER, S.S.; CHUGANI, H.T.; AND SOKOL, R.J. Brain metabolism in children with fetal alcohol syndrome: A positron emission tomography study. Alcoholism: Clinical and Experimental Research 19(Suppl.):53A, 1995.
INGRAM, D.K.; LONDON, E.D.; AND REYNOLDS, M.A. Circadian rhythmicity and sleep: Effects of aging in laboratory animals. Neurobiology of Aging 3:287-297, 1982.
JONES, K.L., AND SMITH, D.W. Recognition of the fetal alcohol syndrome in early infancy. Lancet 2:999-1001, 1973.
KRAJNAK, K.; KASHON, M.L.; ROSEWELL, K.L.; AND WISE, P.M. Aging alters the rhythmic expression of vasoactive intestinal polypeptide mRNA but not arginine vasopressin mRNA in the suprachiasmatic nuclei of female rats. Journal of Neuroscience 18:4767-4774, 1998.
LIANG, F.-Q.; ALLEN, G.; AND EARNEST, D.J. Role of brain-derived neurotrophic factor in the circadian regulation of the suprachiasmatic pacemaker by light. Journal of Neuroscience 20:2978-2987, 2000.
MAIER, S.E.; CHEN, W-J.A.; AND WEST, J.R. The effect of timing and duration of alcohol exposure on development of the fetal brain. In: Abel, E.L., ed. Fetal Alcohol Syndrome. From Mechanisms to Prevention. Boca Raton: CRC Press, 1996. pp. 27-50.
MAIER, S.E.; CRAMER, J.A.; WEST, J.R.; AND SOHRABJI, F. Alcohol exposure during the first two trimesters equivalent alters granule cell number and neurotrophin expression in the developing rat olfactory bulb. Journal of Neurobiology 41:414-423, 1999.
MANTEUFFEL, M.D. Neurotransmitter function: Changes associated with in utero alcohol exposure. In: Abel, E.L., ed. Fetal Alcohol Syndrome. From Mechanisms to Prevention. Boca Raton: CRC Press, 1996. pp. 171-189.
MOORE, R.Y. Disorders of circadian function and the human circadian timing system. In: Klein, D.C.; Moore, R.Y.; and Reppert, S.M., eds. Suprachiasmatic Nucleus: The Mind’s Clock. New York: Oxford University Press, 1991. pp. 429-441.
PICKARD, G.E., AND TUREK, F.W. Effects of partial destruction of the suprachiasmatic nuclei on two circadian parameters: Wheel-running activity and short-day induced testicular regression. Journal of Comparative Physiology 156:803-815, 1985.
PITTENDRIGH, C.S., AND DAAN, S. Circadian oscillations in rodents: A systematic increase of their frequency with age. Science 186:548-550, 1974.
ROEBUCK, T.M.; MATTSON, S.N.; AND RILEY, E.P. Prenatal exposure to alcohol: Effects on brain structure and neuropsychological functioning. In: Hannigan, J.H.; Spear, L.P.; Spear, N.E.; and Goodlett, C.R., eds. Alcohol and Alcoholism: Effects on Brain and Development. Mahwah, N.J.: Lawrence Erlbaum Assoc., 1999. pp. 1-16.
ROOZENDAAL, B.; VAN GOOL, W.A.; SWAAB, D.F.; HOOGENDIJK, J.E.; AND MIRMIRAN, M. Changes in vasopressin cells of the rat suprachiasmatic nucleus with aging. Brain Research 409:259-264, 1987.
ROSETT, H.L.; SNYDER, P.; SANDER, L.W.; ET AL. Effects of maternal drinking on neonatal state regulation. Developmental Medicine and Child Neurology 21:464-473, 1979.
SCHWARTZ, W.J. A clinician’s primer on the circadian clock: Its localization, function and resetting. Advances in Internal Medicine 38:81-106, 1993.
STRATTON, K.; HOWE, C.; AND BATTAGLIA, F., EDS. Fetal Alcohol Syndrome: Diagnosis, Epidemiology, Prevention, and Treatment. Washington, D.C.: National Academic Press, 1996.
SWAAB, D.F.; FISSER, B.; KAMPHORST, W.; AND TROOST, D. The human suprachiasmatic nucleus: Neuropeptide changes in senium and Alzheimer’s disease. Basic and Applied Histochemistry 32:43-54, 1988.
THOMAS, J.D.;
GOODLETT, C.R.; AND WEST, J.R. Alcohol-induced Purkinje cell loss depends
on developmental timing of alcohol exposure and correlates with motor performance.
Developmental Brain Research 105:159-166, 1996.
|
Third Edition of the AOD Thesaurus Is Now Available |
|
The Alcohol and Other Drug Thesaurus: A Guide to Concepts and Terminology in Substance Abuse and Addiction is an invaluable reference guide to the language and nomenclature of the AOD field. Designed for use by researchers and practitioners, the third edition provides information in 4 volumes on more than 10,000 concepts and 2,300 definitions, as well as an extensive network of cross references. A vital indexing tool for information specialists and database managers, the AOD Thesaurus also provides the necessary vocabulary for searching NIAAA’s Alcohol and Alcohol Problems Science Database (ETOH) and for free-text searching other databases. |
|
For information, contact: NIAAA Research Library, c/o CSR, Incorporated, Attn.: Publications Distribution Center, P.O. Box 10686, Rockville, MD 20849–0686. Telephone: (202) 842–7600; fax: (202) 842–0418; Web: http://www.niaaa.nih.gov/publications/thes.htm |
| ||||||||||||||